Okologie und Epidemiologie ¨
Joachim Hermisson 2020
(unter Verwendung von Vorlagen von JH, Ellen Baake und Reinhard B¨urger) 14. November 2020
Literatur
• Sarah P. Otto, Troy Day: A Biologist’s Guide to Mathematical Modeling in Ecology and Evolution, Princeton University Press (∼72 Euro)
• Mark Kot: Elements of Mathematical Ecology, Cambridge University Press (∼62 Euro)
• Nicholas F. Britton: Essential Mathematical Biology, Springer
• Josef Hofbauer, Karl Sigmund: Evolutionary Games and Population Dynamics, Cam- bridge University Press (∼49 Euro)
• Peter Yodzis: Introduction to Theoretical Ecology (1989), Harper & Row.
PDF: www.rug.nl/research/institute-evolutionary-life-sciences/tres/ downloads/bookyodzis.pdf
1
Vorbemerkungen zur Vorlesung WS 2020/2021 2
Biologie und Mathematik
Es ist wohl keine ¨Ubertreibung zu sagen, dass nicht alle großen Biologen talentierte Ma- thematiker waren. Charles Darwin schreibt zu diesem Thema in seiner Autobiographie von 1876:
During the three years which I spent at Cambridge my time was wasted, as far as the academical studies were concerned, as completely as during the three years which I spent at Edinburgh and at school. I attended mathematics, and even went during the summer of 1828 with a private tutor (a very dull man) to Barmouth, but I got on very slowly. The work was repugnant to me, chiefly from my unbeing able to see any meaning in the early steps in algebra. This impatience was very foolish, and in the after-years I have deeply regretted that I did not proceed far enough at least to understand something of the great leading principles of mathematics for men thus endowed seem to have an extra sense. But I do not believe that I should ever have succeeded beyond a very low grade.
Dennoch verdankt Darwin den entscheidenden Durchbruch zu seiner Theorie der nat¨urlichen Selektion einer Kombination von Naturbeobachtung und mathematischer Modellierung, wie er selbst ausdr¨ucklich betont:
In October 1838, that is, fifteen months after I had begun my systematic enqui- ry, I happened to read for amusements ‘Malthus on Population’, and being well prepared to appreciate the struggle for existence which everywhere goes on from long-continued observation of the habits of animals and plants, it at once struck me that under these circumstances favourable variations would tend to be preserved, and unfavourable ones to be destroyed. The result of this would be the formation of new species. Here then I had at last got a theory by which to work.
Die mathematischen Einsichten von Thomas Malthus (1766-1834), dessenEssay on the Prin- ciple of Population (1798) Darwin mehr zuf¨allig in die H¨ande bekam, werden wir im ersten Teil der Vorlesung ausf¨uhrlich besprechen.
Nat¨urlich hat sich seit den Zeiten Darwins in der Biologie viel getan. Generell wird ma- thematische Modellierung ¨uberall dort wichtig, wo die Wissenschaft ¨uber das Sammeln von Daten und Fakten (klassisch: neue Spezies beschreiben, heute: Funktion eines Gens beschrei- ben) hinausgeht und das Gewinnen von neuen Einsichten aus diesen Fakten das Ziel ist. Nach dem Physiker Sir William Bragg ist letzteres die eigentliche Bestimmung jeder Wissenschaft:
The important thing in science is not so much to obtain new facts, but to discover new ways to think about them.
Da die Biologie so komplex und vielf¨altig ist wie keine andere Naturwissenschaft, steht h¨aufig noch das Sammeln von Daten und Fakten an erster Stelle. Zunehmend k¨onnen heute aber immer gr¨oßere Mengen an Daten in kurzer Zeit gewonnen werden (Sequenzdaten, Genexpres- sionen), sodass nicht das Sammeln selbst, sondern die Gewinnung neuer Erkenntnisse aus diesen Daten zum vorrangigen Ziel wird. Dies erfordert oft komplexe Modelle – eine zuneh- mende Mathematisierung der Wissenschaft ist die Folge.
Es wird in dieser Vorlesung nicht m¨oglich sein, einen umfassenden ¨Uberblick ¨uber Model- le und Methoden der mathematischeen Biologie zu geben. Das Ziel ist es vielmehr, eine Idee
Vorbemerkungen zur Vorlesung WS 2020/2021 3 davon zu vermitteln, was es heißt “mathematisch zu modellieren” und mit welchen Methoden man in Modellen zu biologischem Verst¨andnis kommen kann. Dies wird im ersten Teil der Vorlesung beispielhaft anhand einiger Modelle aus der ¨Okologie und der Epidemiologie ge- schehen. Im zweiten Teil werden Modelle aus der Populationsgenetik und der (evolution¨aren) Spieltheorie behandelt.
Mathematische Modelle in der Biologie
Jedes biologische Modell ist eine Abbildung eines Teils der Natur auf einen mathematischen Formalismus. Modelle sind immer Abstraktionen, d.h. vereinfachende Darstellungen der Wirk- lichkeit. Modellierung beginnt deshalb mit einer Reihe von Modellannahmen: Einige Aspekte eines in der Natur beobachteten Systems werden in das Modell integriert, weil wir davon ausgehen, dass sie f¨ur das vorliegende Problem wesentlich sind. Viele andere Aspekte werden ignoriert (bzw. es wird von ihnen abstrahiert), entweder weil sie wahrscheinlich unwichtig sind oder weil wir bewußt eine reduktionistische Perspektive einnehmen wollen. Im letzteren Fall hoffen wir, dass wir ein komplexes System verstehen k¨onnen, indem wir (Gruppen von einzel- nen) Faktoren nacheinander untersuchen. Wenn wir z.B. die zuk¨unftige Bev¨olkerungsgr¨oße in Osterreich modellieren, sind die derzeitige Gr¨¨ oße und die Altersstruktur sicherlich wesentlich.
Andere Faktoren wie Fortschritte in der medizinischen Behandlung k¨onnten ebenfalls einen gewissen Einfluss auf die Sterblichkeitsraten haben, k¨onnen aber in einem einfachen Modell ignoriert werden. Noch andere Faktoren, wie Immigration, sind wahrscheinlich wichtig, aber eine Behandlung ohne Einwanderung k¨onnte uns bereits wertvolle Informationen liefern bevor wir die Auswirkungen von Immigration in einem separaten Schritt untersuchen.
Je mehr Faktoren in ein Modell einbezogen werden, desto pr¨aziser und spezifischer wird es. Dies ist insbesondere f¨ur zuverl¨assige quantitative Vorhersagen erforderlich (Wettervor- hersage, demographische Modelle). Gr¨oßere Komplexit¨at bedeutet aber immer auch weniger Handhabbarkeit und meist auch weniger Allgemeing¨ultigkeit. Aus einem Modell, das genauso komplex ist wie das System, das es repr¨asentiert, k¨onnen wir keine neuen Erkenntnisse gewin- nen. Komplexe quantitative Modelle, die f¨ur Vorhersagen verwendet werden, k¨onnen in der Regel nur durch Computersimulationen behandelt werden. Im Gegensatz dazu sind viele Fra- gen qualitativer Natur, ob zwei Arten koexistieren k¨onnen oder wann es zu einem Ausbruch einer Infektionskrankheit kommen kann. In diesen F¨allen ist es das Ziel der Modellierung, einen minimalen Satz von Faktoren zur Erkl¨arung eines Ph¨anomens zu finden. Ein Modell ist dann umso besser, je einfacher es ist.
Die Kunst der Modellierung besteht also darin, die wesentlichen Faktoren auszuw¨ahlen, die in ein Modell einfließen sollen. Dies erfordert einerseits eine gewisse Erfahrung mit dem zugrundeliegenden (z.B. biologischen) System. Andererseits erfordert es Verst¨andnis des ma- thematischen Mechanismus, um zu erkennen, welche Faktoren entscheidende Konsequenzen haben k¨onnen, auch wenn ihre Effekte zun¨achst klein aussehen. Zu diesem Zweck st¨utzt sich die Modellierung auf einen breiten mathematischen Werkzeugkasten, der Elemente aus der Theorie stochastischer Prozesse, dynamischer Systeme, Differentialgleichungen und der Sta- tistik enth¨alt.
Auch wenn die Motivation f¨ur eine ernsthafte Modellbildung in ihrer Anwendung liegt, tr¨agt sie auch in der Mathematik als eigenst¨andiger Wissenschaft Fr¨uchte. Oft zeigen sich in der Anwendung neue Strukturen, die mit den verf¨ugbaren mathematischen Methoden nur unzureichend beschrieben werden k¨onnen. Historisch betrachtet gehen deshalb viele der be- deutendsten Entwicklungen in der Mathematik, wie die Differential- und Integralrechnung
Vorbemerkungen zur Vorlesung WS 2020/2021 4 oder die Wahrscheinlichkeitstheorie, auf ganz konkrete Probleme in der Anwendung zur¨uck.
Neben der Physik hat auch die Biologie immer wieder hierzu beigetragen, etwa bei der Ent- wicklung der mathematischen Statistik durch R.A. Fisher und durch wesentliche Impulse f¨ur die Theorie dynamischer Systeme und stochastischer Prozesse.
Okologie und Epidemiologie¨
Oikos= Haus, Wohnung.Logos = Wort, Studium von. ¨Okologie bezieht sich auf die wissen- schaftliche Untersuchung lebender Organismen in ihrer nat¨urlichen Umgebung. Sie ist eine vielf¨altige wissenschaftliche Disziplin und deckt verschiedene Ebenen der biologischen Orga- nisation ab.
• Auf der individuellen Ebene diskutiert diephysiologische ¨Okologieden Einfluss von Nah- rung, Licht, Feuchtigkeit, Pestizidkonzentrationen usw. auf das Leben von Individuen.
• Die Populations¨okologie untersucht die Wechselwirkungen von Populationen mit ihrer Umwelt und ihre Auswirkungen auf die Bev¨olkerungsstruktur und Demographie. Auf der gleichen Ebene diskutiert dieVerhaltens¨okologiedie Konsequenzen unterschiedlicher Verhaltensstrategien.
• Schließlich behandeln dieGemeinschafts¨okologieund dieOkosystemforschung¨ das Schick- sal komplexer ¨Okosysteme mit zwei bis zehntausenden interagierender Arten und Ar- tengruppen.
Die Wechselwirkungen von Populationsdynamik und evolution¨arer Dynamik sind Gegenstand derevolution¨aren ¨Okologie. Viele Zweige der ¨okologischen Forschung verwenden mathemati- sche Modelle. So bedient sich die Verhaltens¨okologie beispielsweise spieltheoretischer Metho- den, um die Auswirkungen von Verhaltensstrategien zu untersuchen. Die Evolutions¨okologie st¨utzt sich stark auf die mathematischen Modelle der Evolutionsgenetik. Das Thema dieser Vorlesung muss enger gefasst werden und wird sich haupts¨achlich auf die Populations¨okologie konzentrieren, wo wir die Dynamik von Populationsgr¨oßen und ihre Gleichgewichte untersu- chen. Als wesentliche Anwendung werden wir in einem zweiten Teil dann Modelle der Epi- demiologie besprechen, die die Dynamik von infekti¨osen Krankheiten in einer Population beschreiben.
Arbeitsblatt 1 zur Vorlesung WS 2020/2021 5
1 Elementare Populationsdynamik in einer Dimension
Populationsgr¨oßen biologischer Arten und ihre zeitliche Entwicklung geh¨oren zu den funda- mentalsten Bestimmungsgr¨oßen der ¨Okologie. Mathematisch ist der dynamische Prozess des Populationswachstums und -r¨uckgangs ganz allgemein eine Funktion von Faktoren, die einer Population immanent sind (z.B. ihr Reproduktionspotenzial, ihr Lebenszyklus oder ihre Dich- te) und von Umweltbedingungen. Die Umwelt umfasst alle Ressourcen, die f¨ur das Gedeihen einer Population unerl¨asslich sind, wie Nahrung und Raum, sowie Faktoren, die ihre Gr¨oße verringern k¨onnen, wie Raubtiere und Krankheiten. In der Natur sind viele dieser Faktoren in der Tat selbst dynamische Populationen, die als Nahrungsquelle, Konkurrent, Raubtier oder Parasit eine direkte Auswirkung auf die Dynamik der fokalen Population haben. Da die dyna- mischen Prozesse von (z.B.) Raubtieren und Beutetieren interagieren, erhalten wir schnell ein komplexes mehrdimensionales Problem. Mit diesen Komplexit¨aten werden wir uns zweiten und dritten Kapitel der Vorlesung ausf¨uhrlich besch¨aftigen. In einem ersten Schritt gehen wir jedoch vereinfachend davon aus, dass wir alle Wechselwirkungen mit anderen dynami- schen Aspekten der Umwelt ignorieren k¨onnen und modellieren die Dynamik einer einzelnen isolierten Population. Biologisch ist solch eine vereinfachte Betrachtung manchmal gerechtfer- tigt, zum Beispiel wenn sich die Dynamik der interagierenden Populationen auf verschiedenen Zeitskalen abspielt: entweder viel schneller, so dass wir davon ausgehen k¨onnen, dass sich die interagierende Population in einem dynamischen Gleichgewicht befindet, oder viel langsamer, so dass sich die Gr¨oße einer interagierenden Population ¨uber die interessierenden Zeitspannen nicht wesentlich ¨andert. Als weitere wesentliche Annahme gehen wir davon aus, dass die Po- pulation unstrukturiert ist. Dies bedeutet, dass alle Individuen der Population als gleichwertig behandelt werden. Insbesondere gibt es keine Altersklassen, keine ph¨anotypischen Unterschie- de (die f¨ur die Dynamik von Bedeutung sind), und wir k¨onnen die Verteilung der Population
¨
uber den physischen Raum ignorieren.
1.1 Deterministische Prozesse in diskreter Zeit
Viele Prozesse in der Natur werden als in der Zeit kontinuierlich ablaufend betrachtet. F¨ur andere, wie beispielsweise das Wachstum einer Insektenpopulation mit getrennten Genera- tionen oder f¨ur Kreuzungsexperimente, ist eine diskrete Zeitskala sinnvoller, auf der nur be- stimmte Zeitpunkte, z.B. Vielfache der Generationsdauer ∆t, angegeben werden. Eine diskrete Betrachtungsweise entspricht oft auch der experimentellen Situation, wo meist nicht konti- nuierlich, sondern in regelm¨aßigen Abst¨anden gemessen wird. Messen wir beispielsweise die Populationsgr¨oßex zu den Zeitpunkten 0,∆t,2∆t,3∆t, . . ., so bilden die Werte x0, x1, x2. . . eine Folge, die die zeitliche Ver¨anderung des Systems beschreibt. Eine Folge ist iterativ (oder rekursiv) definiert, wenn eine Vorschrift f gegeben ist, wie man ein Folgenglied aus dem jeweils vorhergehenden gewinnt:
xn+1=f(xn). (1)
Die Iterationsfunktion f wird im populationsbiologischen Zusammenhang auch als Repro- duktionsfunktion bezeichnet. Sie stellt eine dynamische Regel dar, die angibt, wie sich die Populationsgr¨oße von einem Zeitschritt (Generation, Jahr) zum n¨achsten ver¨andert. Die zu- geh¨orige Folge heißt auch Zeitverlauf oder Populationsentwicklung.
Der entscheidende Schritt zur mathematische Modellierung eines biologischen Prozesses ist die Konstruktion der Reproduktionsfunktion aus biologisch motivierten ¨Uberlegungen ¨uber das System, das man beschreiben m¨ochte. Im folgenden werden wir einige Beispiele hierf¨ur
Arbeitsblatt 1 zur Vorlesung WS 2020/2021 6 kennenlernen. In einem zweiten Schritt m¨ochte man dann wissen, wie die Reproduktionsfunk- tion den Zeitverlauf des biologischen Prozesses bestimmt.
Modellbildung I: Geometrisches Wachstum
Betrachten wir, um konkret zu werden, die Vogelpopulation auf einer Insel. Jedes Jahr wird die Populationsgr¨oße bestimmt. Wie kann man die Entwicklung dieser Gr¨oße in einem Modell beschreiben? F¨ur das einfachste Modell machen wir die folgenden Annahmen:
Als erstes nehmen wir an, dass die Populationsgr¨oße im Folgejahr,xn+1, nur von der aktu- ellen Populationsgr¨oßexn abh¨angt, und nicht von anderen in der Zeit ver¨anderlichen Gr¨oßen (wir vernachl¨assigen also z.B. Klimaschwankungen oder den Einfluß von Populationsgr¨oßen anderer Vogelarten). Außerdem nehmen wir an, dass sich die Populationsgr¨oße zwischen den Jahern nur durch Geburten und Todesf¨alle ¨andert. Wir setzen also anxn+1 =xn+Bn−Dn, wobei Bn und Dn die Anzahl der Geburten und Todesf¨alle in einer Population der Gr¨oße xn angibt. In der einfachsten N¨aherung sollten diese Zahlen einfach proportional zur ur- spr¨unglichen Zahl der Individuen sein, alsoBn=bxnundDn=dxn.bunddsind Konstanten und geben die mittlere Zahl der Geburten und Todesf¨alle in der Population pro Individuum und pro Jahr an. Mit der Definitionr:= 1 +b−dfinden wir damit die Reproduktionsfunktion xn+1=f(xn) =xn+bxn−dxn=rxn. (2) Die Reproduktionsfunktionf(x) =rxist einfach eine Gerade durch den Ursprung (s. Abb. 3).
F¨ur jeden Anfangswert x0 k¨onnen wir durch wiederholtes Anwenden der Iterationsvorschrift (2) die Entwicklung der Populationsgr¨oße in aufeinanderfolgenden Jahren voraussagen, d.h:
x1 =rx0, x2 =rx1 =r2x0, ...
Offensichtlich l¨aßt sich der Zeitverlaufexplizitangeben als geometrische Folge
xn=rnx0. (3)
0 2 4 6 8
Zeit 0
250 500 750 1000 1250 1500
Population
0 2 4 6 8
Zeit 2.3
3.3 4.3 5.3 6.3 7.3
Lnxn
0 200 400 600 800
Population jetzt 0
250 500 750 1000 1250 1500
Population1Jahrspaeter
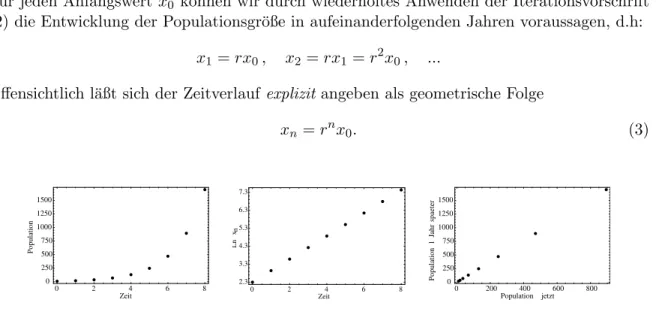
Abbildung 1: Geometrisches Wachstum einer Vogelpopulation. Links: Populationsgr¨oße xn
als Funktion der Zeit n in Jahren; Mitte: Logarithmus der Populationsgr¨oße (sog. halb- logarithmische Auftragung); rechts: Populationsgr¨oße in der n¨achsten Generation als Funktion der aktuellen Gr¨oße (die Punkte liegen auf dem Graphen der Reproduktionsfunktion).
Beispiel In einer Vogelpopulation, die eine Insel neu besiedelt hat, werden pro Jahr und Individuum im Mittel 1.1 neue V¨ogel geboren und 0.2 sterben. Das f¨uhrt zu geometrischem Wachstum mit r= 1.9, alsoxn=rnx0, mitx0 = 10 in Zahlen:
Arbeitsblatt 1 zur Vorlesung WS 2020/2021 7
Zeit n(in Jahren) 0 1 2 3 4 5 6 7 8
Populationsgr¨oße xn 10 19 36 69 130 248 470 894 1698 Betrachtet man anstelle der Populationsgr¨oße den (nat¨urlichen) Logarithmus,
yn= ln(xn) = ln(rnx0) =nln(r) + ln(x0),
so erh¨alt man einen linearen Anstieg mit Steigung ln(r) und Achsenabschnitt ist ln(x0). An dieser Darstellung l¨aßt sich das geometrische Wachstum besonders leicht erkennen. Allgemein erhalten wir f¨ur eine Reproduktionsfunktion der Form f(x) =rx ein qualitativ unterschied- liches Verhalten abh¨angig vom Wert vonr≥0:
r= 0 sofortiges Aussterben, 0< r <1 geometrisches “Schrumpfen”,
r= 1 xn=x0 f¨ur alle n, r >1 geometrisches Wachstum . Modellbildung II: Ein komplexeres Modell
Wir betrachten wieder das Beispiel der Vogelpopulation auf einer Insel. Im letzten Abschnitt hatten wir mit der Iterationsfunktionf(x) = (1 +b−d)x=rx ein sehr einfaches Modell f¨ur die zeitliche Dynamik der Populationsgr¨oße konstruiert. Dieses Modell ent¨alt zwei Parameter, die Geburtenratebund die Todesrated, die wir aus Beobachtungen im Feld sch¨atzen k¨onnen (die Dynamik h¨angt allerdings nur von der Differenzb−d ab). Um das Modell des geome- trischen Wachstums realistischer zu machen, wollen wir nun einige Aspekte, die wir bisher vernachl¨assigt hatten, erg¨anzen:
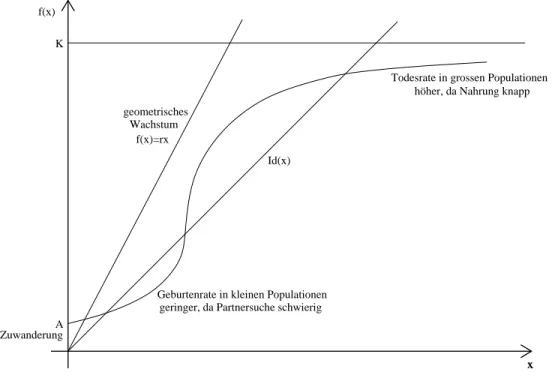
• Jedes Jahr wandern einige V¨ogel vom Festland auf die Insel zu. Wir modellieren dies durch einen Beitrag A zu f(x), wobei Aeinfach eine positive Zahl unabh¨angig von der Populationsgr¨oße xist.
• Bisher haben wir eine konstante Geburtenrate b angenommen. Die Zahl der Neugebo- renen ist dann einfach proportional zur Populationsgr¨oße B(x) = bx. F¨ur sehr kleine Populationen ist das nicht realistisch. Der Grund ist, dass zur Produktion von Nachkom- men erst einmal ein Partner gefunden werden muss. Dies gelingt bei kleiner Populati- onsgr¨oßexnicht immer. Die effektive Anzahl der Geburten ist deshalb zun¨achst kleiner und erreicht erst f¨ur ein hinreichend großes x (wenn die Partnersuche kein Problem mehr ist) die RelationB(x) =bx. Biologisch nennt man dies einenAlle-Effekt.
• Eine weitere unrealistische Eigenschaft des geometrischen Wachtums-Modells ist es, dass Populationen unendlich groß werden k¨onnen. In Wirklichkeit ist die Populationsgr¨oße durch den beschr¨ankten Vorrat an Nahrung und Platz immer begrenzt. Wir koennen dies in unserem Modell durch eine maximale Populationsgr¨oßeK (die sogenanntecar- rying capacity) ber¨ucksichtigen. Die Iterationsfunktion f(x) kann den Wert K nicht ubersteigen, sondern n¨¨ ahert sich ihm f¨ur große xasymptotisch an.
Die ¨Anderungen, die sich aus diesen ¨Uberlegungen f¨ur die Reproduktionsfunktionf(x) erge- ben sind in der Abbildung 2 dargestellt.
Arbeitsblatt 1 zur Vorlesung WS 2020/2021 8
x A
K
x f(x)
geometrisches
Zuwanderung
Todesrate in grossen Populationen
Wachstum f(x)=rx
höher, da Nahrung knapp
geringer, da Partnersuche schwierig Geburtenrate in kleinen Populationen
Id(x)
Abbildung 2: Reproduktionsfunktionen f¨ur das geometrische und das komplexere Modell.
Modellanalyse I: Cobwebbing
Eine graphische Methode, die es erlaubt, auch f¨ur kompliziertere Reproduktionsfunktionen qualitative Aussagen ¨uber die Entwicklung der Populationsgr¨oße zu treffen ist das sogenannte Cobwebbing:
1. Zun¨achst zeichnen wir den Graphen der Reproduktionsfunktionf und den Graphen der Winkelhalbierenden Id(x) =x und w¨ahlen den Anfangswertx0.
2. Umx1 ausx0 zu bestimmen, wenden wirf aufx0 an, d.h.x1 ist der Funktionswert von f an der Stellex0. Wir zeichnen einen Pfeil von Punkt (x0, x0) zum Punkt (x0, x1) auf dem Graphen von f.
3. Um x2 ausx1 zu bestimmen, m¨ussen wir zun¨achst den Wert x1 von der vertikalen auf die horizontale Achse ¨ubertragen. Dies erreichen wir dadurch, dass wir einen waage- rechten Pfeil vom Punkt (x0, x1) auf dem Graphen von f zum Punkt (x1, x1) auf der Winkelhalbierenden zeichnen. Nun erh¨alt man x2 = f(x1) analog zu Schritt 2 durch einen senkrechten Pfeil von (x1, x1) zum Punkt (x1, x2) auf dem Graphen vonf. Wenn man Schritt 3 von verschiedenen Anfangspunkten x0 aus wiederholt ausf¨uhrt, erh¨alt man das in Abb. 3 dargestellte Bild.
Gleichgewichtspunkte
Von besonderem Interesse in einem dynamischen Prozess sind die Werte x∗, auf die sich die dynamische Variable (z.B. die Populationsgr¨oße) einstellt wenn man lang genug wartet, also limn→∞xn=x∗. Wir definieren:
Arbeitsblatt 1 zur Vorlesung WS 2020/2021 9
x Id(x)
f(x)
Abbildung 3: Iterationsfunktion und Cobwebbing f¨ur das Modell einer Vogelpopulation. Die Reproduktionsfunktion selbst gibt nicht die Populationsgr¨oße als Funktion der Zeit an. Die Zeitentwicklung gewinnen wir erst durch das Cobwebbing.
Definition: Gleichgewichts- oder Fixpunkt Sei f eine Reproduktionsfunktion. Dann heißt eine L¨osung x∗ der Gleichung
f(x) =x (4)
Fixpunkt oder Gleichgewichtspunkt. Istx∗ ein Fixpunkt undxn=x∗, dann istxn+1 =xn= x∗. Graphisch lassen sich die Fixpunkte bestimmen als Schnittpunkte der Reproduktionsfunk- tion mit der Winkelhalbierenden. Wichtig f¨ur die Analyse des dynamischen Prozesses ist nun die Unterscheidung von stabilen und instabilen Fixpunkten:
• Ein Fixpunkt x∗ heißt global stabil, wenn die Folge der xn unabh¨angig vom Startwert x0 gegen x∗ konvergiert.
• Ein Fixpunkt x∗ heißt lokal stabil, wenn die Folge der xn f¨ur solche x0, die nah genug an x∗ liegen, gegenx∗ konvergiert.
• Ein Fixpunkt x∗ heißt instabil, wenn er nicht lokal stabil ist.
Kriterium f¨ur (lokale) Stabilit¨at bzw. Instabilit¨at: Ob ein Fixpunkt stabil oder insta- bil ist, k¨onnen wir am konkreten Beispiel meist leicht mit dem Cobwebbing herausfinden. F¨ur die Vogel-Population erhalten wir zum Beispiel zwei stabile und einen instabilen Fixpunkt (siehe Abb. 3). Mit Hilfe des graphischen Verfahrens k¨onnen wir aber auch ein allgemeines Kriterium f¨ur die lokale Stabilit¨at eines Fixpunktes ermitteln. Wir betrachten dazu wie sich das Cobweb in Abh¨angigkeit der Steigung der Reproduktionsfunktion am Schnittpunkt mit der Winkelhalbierenden ¨andert. Wir sehen (vgl. Abb. 3):
Arbeitsblatt 1 zur Vorlesung WS 2020/2021 10
• Ist die Steigung der Reproduktionsfunktion am Fixpunkt gr¨oßer als 1, f0(x∗)>1 (f(x) schneidet die Winkelhalbierende von unten nach oben), so kommt man auf dem Cobweb immer weiter vom Fixpunkt weg: der Fixpunkt ist instabil.
• Ist die Steigung 0≤f0(x∗)<1, so n¨ahert man sich auf dem Cobweb dem Fixpunkt von beiden Seiten aus an: der Fixpunkt ist (lokal) stabil. Dabei ¨uberspringt die Folgexndas Gleichgewicht nicht, sondern bleibt jeweils auf einer Seite: monotone Konvergenz.
• F¨ur−1< f0(x∗)<0 ist der Fixpunkt ebenfalls lokal stabil, aber die Folgexn¨uberspringt das Gleichgewicht mit jedem Schritt. Wir habenoszillierende Konvergenz (oder ged¨ampfte Schwingungen).
• Ist die schließlich f0(x∗) < −1, so ist der Fixpunkt wiederum instabil (oszillierende Divergenz).
• Ist die Steigung genau gleich eins oder gleich−1, so l¨aßt dieses Kriterium keine Aussage zu. Das Kriterium liefert auch keine Information ¨uber das globaleVerhalten.
Ubungsaufgaben 1¨
Aufgabe 1.1: Ein Baum w¨achst jedes Jahr um α cm.
(a) Geben Sie die Iterationsfunktion an und stellen Sie sie graphisch dar und f¨uhren Sie das Cobwebbing f¨ur ein beliebig gew¨ahltes x0 durch.
(b) Geben Sie die Baumh¨ohe im n-ten Jahr explizit an. Um welche Folge handelt es sich?
Aufgabe 1.2: Radioaktive Isotope, wie C14, zerfallen im Laufe der Zeit. Je nach Isotop zerf¨allt jedes Jahr ein fester Anteil des noch vorhandenen Materials. Man charakterisiert Iso- tope durch ihreHalbwertszeit, das ist der Zeitraum, innerhalb dessen die H¨alfte des Materials zerf¨allt. Die Halbwertszeit von C14 ist 5730 Jahre.
(a) Geben Sie die Iterationsfunktion an und ermitteln Sie, wieviel Prozent des noch vor- handenen Materials pro Jahr zerf¨allt.
(b) Das Mischungsverh¨altnis des radioaktiven C14und des nichtradioaktiven C12im CO2der Atmosph¨are ist ¨uber geologische Zeitskalen konstant geblieben, und Pflanzen bauen es in demselben atmosph¨arischen Mischungsverh¨altnis in ihre organischen Molek¨ule (z.B.
Cellulose, Lignin) ein. Wenn die Pflanze stirbt, nimmt sie kein neues CO2 mehr auf.
Das radioaktive C14 in dem toten Pflanzenmaterial zerf¨allt (gem¨aß der uns bekannten Iterationsfunktion), das C12bleibt unver¨andert. Somit sinkt das Verh¨altnis C14/C12 im Laufe der Zeit ab. Geben Sie die Iterationsfunktion f¨ur dieses Verh¨altnis an.
(c) Bei einer Ausgrabung wird ein alter Holzrest gefunden, dessen C14/C12 Verh¨altnis nur noch 23 % desjenigen Wertes betr¨agt, den man in lebendem Pflanzenmaterial findet.
Finden Sie heraus, wann das Holz geschlagen wurde. Benutzen Sie dazu die vorher ermittelte Zerfallsrate.
Arbeitsblatt 1 zur Vorlesung WS 2020/2021 11 Aufgabe 1.3: In zwei Beh¨altern I und II befinden sich insgesamt γ Mol einer gel¨osten Substanz. Die Beh¨alter sind durch eine Membran getrennt, die eine gewisse Durchl¨assigkeit f¨ur die Partikel besitzt. Die Beweglichkeit der Teilchen, die Durchl¨assigkeit der Membran und die Gr¨oße bzw. Form der Beh¨alter sollen so sein, daß jeweils nach 1 min. ein Anteilαder zuvor in I befindlichen Teilchen nach II gewandert ist und ein Anteilβ der zuvor in II befindlichen nach I. Wenn xn die Stoffmenge (in Mol) in I und yn die Stoffmenge in II nach n Minuten ist, dann gilt
xn+1 =xn−αxn+βyn, xn+yn=γ wobei 0< α+β <1 sein soll.
(a) Berechnen Sie die resultierende Iterationsfunktion f¨ur die Stoffmenge in I, bestimmen Sie den Fixpunkt und die Stabilit¨at.
(b) Setzen Sie nunγ = 0.2,α = 0.2 undβ = 0.1. Ermitteln Sie (graphisch oder rechnerisch) f¨ur die ersten 5 Minuten die Zeitentwicklung f¨ur die Stoffmenge in I, und zwar f¨ur die Startwerte x0 = 0 und x0 = 0.15. Zeichnen Sie beide Zeitverl¨aufe in ein Bild. Auf welchen Wert werden sich die Stoffmengen in I und II einstellen?
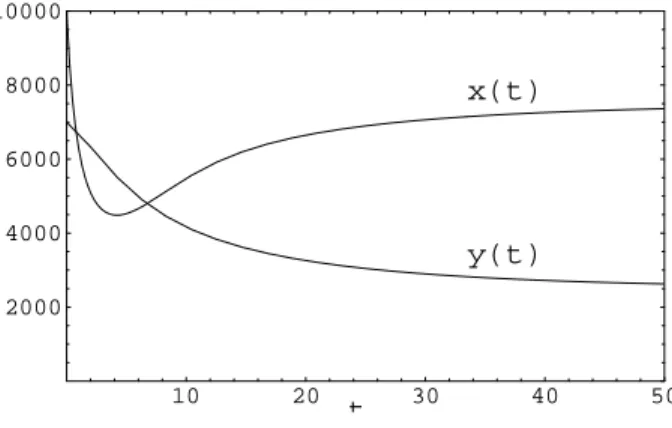
Aufgabe 1.4: Ein Bienenvolk verliert w¨ahrend der Sommermonate in jeder Woche 20% der zu Beginn dieser Woche vorhandenen Arbeiterinnen (weil diese sterben oder nicht zum Stock zur¨uckfinden). Es schl¨upfen aber jede Woche 7000 neue Arbeiterinnen.
(a) Wie lautet die Iterationsfunktion? Zeichnen Sie diese Iterationsfunktion. Gibt es Fix- punkte? Sind diese global oder lokal stabil?
(b) K¨onnen Sie eine analoge Iterationsfunktion f¨ur die Zahl der Arbeiterinnen nach zwei Wochen angeben, also f¨ur xn+2 als Funktion von xn? Vergleichen Sie mit der Iterati- onsfunktion f¨ur eine Woche.
Aufgabe 1.5: Schauen Sie sich das Cobwebbing f¨ur unterschiedliche Iterationsfunktionen an und versuchen Sie hieraus Kriterien f¨ur die globale Stabilit¨at von Fixpunkten zu formulieren.
Hinweis: Das geht im allgemeinen Fall nicht so eindeutig wie f¨ur die lokale Stabilit¨at. Man kann aber Kriterien f¨ur große Klassen von Iterationsfunktionen finden (f hat einen global stabilen Fixpunkt wenn . . . ).
Arbeitsblatt 2 zur Vorlesung WS 2020/2021 12 1.2 Kontinuierliche Entwicklungsprozesse
Bislang haben wir Prozesse in diskreter Zeit untersucht, d.h. wir haben angenommen, daß nur zu festen Zeitpunkten etwas passiert, z.B. daß sich Bakterien alle gleichzeitig einmal pro Stun- de teilen. Eine solche Annahme ist jedoch nicht immer gerechtfertigt; oft ist es realistischer, anzunehmen, daß der Zuwachs kontinuierlich in der Zeit stattfindet. Wir werden jetzt also eine kontinuierliche Zeitvariablet ≥0 betrachten, die wir auf einer beliebigen Skala messen (Sekunden, Jahre, ...).N(t) bezeichnet die (mittlere) Populationsgr¨oße oder auch die Dichte einer einer Population, z.B. die Zahl der Individuen pro Quadratmeter und kann ebenfalls kontinuierliche Werte inR+ annehmen. Sei ∆tein kurzes Zeitintervall. Wir nehmen an, dass mit einer konstanten Geburtenrate b (pro Individuum) Nachkommen geboren werden und dass Individuen mit einer Todesrate dsterben. Im einfachsten Modell sind diese Raten zeit- lich konstant und f¨ur alle Individuen in der Population gleich. Insbesondere h¨angen sie nicht vom Alter ab. Dann ergibt sich die ¨Anderung der Populationsgr¨oße im Zeitintervall ∆t zu
∆N(t) =N(t+ ∆t)−N(t) = (b−d)∆tN(t) (5) Im Grenzfall ∆t→0 ergibt sich
N˙(t) = lim
∆t→0
∆N(t)
∆t = (b−d)N(t) =λN(t) (6)
wobei λ = b−d die sogenannte (intrinsische) Wachstumsrate ist (oder: der Malthus’sche Parameter nach Thomas Malthus, 1766-1834, der das Modell als erster auf die menschliche Population angewendet hat). Die eindeutige L¨osung dieser gew¨ohnlichen Differentialgleichung mit AnfangsbedingungN(0) =N0 ist
N(t) = exp[λt]N0. (7)
Exponentielles Wachstum in kontinuierlicher Zeit ist das Gegenst¨uck zum geometrischen Wachstum in diskreter Zeit. Die enge Beziehung sieht man auch aus Gleichung (7) wenn man die Konstante exp[λ] als Wachstumsparameter r w¨ahlt. Dann ist
N(t) = exp[λ]t
N0 =rtN0.
Wenn man nun den in kontinuierlicher Zeit ablaufenden Prozess in diskreten Zeitschritten beschreibt (weil man etwa einmal in der Stunde misst), erh¨alt man wieder ein geometrisches Wachstum. In diesem Sinne sind exponentielles und geometrisches Wachstum “ein und dassel- be”. W¨ahrend beim geometrischen Wachstum die Grenze zwischen Schrumpfen und Wachsen bei r = 1 verl¨auft, liegt sie beim exponentiellen Wachstum bei λ = 0. Mit λ > 0 folgt Bev¨olkerungsexplosion und ibei λ <0 stirbt die Population aus.
Das allgemeine deterministische Modell f¨ur die Dynamik einer einzelnen, unstrukturierten Population lautet:
N˙(t) =f(N, t) =c(t) +λ N(t), t
N(t). (8)
Dabei beschreibtc(t) die Immigration von Individuen undλ N(t), t
eine allgemeine Wachs- tumsrate pro Kopf. Abgesehen vom exponentiellen Wachstum gibt es nur wenige Modelle mit einer expliziten L¨osung.
Arbeitsblatt 2 zur Vorlesung WS 2020/2021 13
• Das einfachste Modell mit einer stabilen Gleichgewichtspopulation N(t)>0 ergibt sich aus einer Dynamik mit Einwanderung und Tod (negative Wachstumsrate),
N˙(t) =f(N) =c−dN(t) (9)
mitc, d >0 und L¨osung
N(t) = c d+
N0−c
d
exp[−dt]. (10)
In der Populationsdynamik beschreibt das Modell eine Populationssenke mit Immigra- tion und Tod. Alternativ beschreibt es in einem Kontinent-Insel Modell die ¨Anderung einer lokalen Populationsdichte auf der “Insel” (oder einem anderen lokalen Habitat) durch konstante Einwanderung von einem “Kontinent” (einem Reservoir mit konstan- ter Populationsdichte) und Auswanderung mit einer konstanten Rate pro Kopf. In der Biochemie entspricht dies dem Konzentrationsausgleich durch Diffusion eines Stoffes durch eine Zellmembran, wobei N(t) in dieser Interpretation die variable Konzentrati- on des Stoffes in der Zelle ist und die Konzentration außerhalb der Zelle als konstant angenommen wird.
• Das einfachste Modell mit Populationsregulierung in einer Population, die sich selbst erhalten kann, ist das logistische Wachstum mit
N˙(t) =λN(t)
1− N(t) K
. (11)
Das Modell ist durch eine lineare Dichteabh¨angigkeit mit einer Pro-Kopf-Wachstumsrate von λ(1−N/K) gekennzeichnet. Der Parameter λ ist die intrinsische Wachstumsrate in Abwesenheit von innerartlicher Konkurrenz. K ist die sogenanntecarrying capacity (Tragf¨ahigkeit). F¨ur N(t) > K ist die Konkurrenz so groß (die Ressourcen werden so knapp), dass die Population schrumpft. Die explizite L¨osung f¨ur das logistische Wachs- tumsmodell folgt (z.B. durch Trennung der Variablen) als
N(t) = K
1−(1−K/N0) exp[−λt]. (12)
Das logistische Wachstum kann auch f¨ur zeitabh¨angige Pro-Kopf-Wachstumsrate λ(t) noch explizit gel¨ost werden, wobei man einfach den ExponentλtdurchRt
0λ(t)dtersetzt.
Die allerwenigsten Modelle in der theoretischen ¨Okologie haben explizite L¨osungen. Gl¨uck- licherweise sind wir in der Regel nicht so sehr an der expliziten Dynamik interessiert, sondern eher an den qualitativen Eigenschaften wie der Gleichgewichtsstruktur.
Modellanalyse II: Phasenliniendiagramm
Analog zum Cobwebbing im diskreten Fall kann man die Zeitentwicklung auch f¨ur komplizierte Differentialgleichungen graphisch analysieren. Wir betrachten dazu eine beliebige autonome Differentialgleichung in einer Dimension,
N˙(t) =f(N), (13)
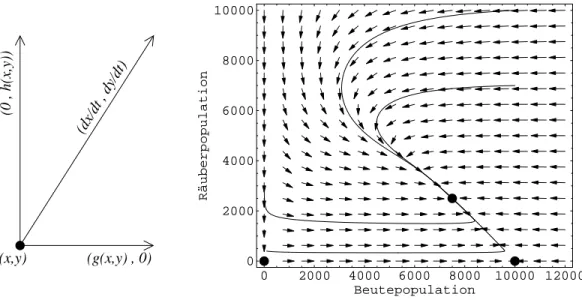
bei der die sogenannte Geschwindigkeitsfunktionf(N) nur indirekt ¨uber N =N(t) von der Zeit abh¨angt. All diese Systems kann man mit der Methode desPhasenliniendiagrammsquali- tativ untersuchen, das in Abbildung 4) f¨ur das Beispiel des Migrations- oder Diffusionsmodells dargestellt ist.
Arbeitsblatt 2 zur Vorlesung WS 2020/2021 14
N f(N)
c/d c
0 5 10 15 20
tHin JahrenL 2
4 6 8 10 12 14 16
NHtL
Abbildung 4: Migrationsmodell gem¨aß Gleichung (9). Links: Phasenliniendiagramm. Rechts:
Die zugeh¨orige Zeitentwicklung N(t) f¨ur c = 1, d = 0.1 (pro Jahr) und zwei verschiedene Anfangsdichten N0 = 3 undN0= 15.
1. Zeichne den Graphen der Funktionf(N) und denkeN(t) als Bewegung auf derx-Achse, deren Geschwindigkeit ˙N an jedem Punkt N durch den Wert f(N) gegeben ist.
2. Deute die Entwicklung der dynamischen Gr¨oße in jedem Punkt N durch Pfeile auf der x-Achse an: Wenn ˙N = f(N) positiv (negativ) ist, wirdN gr¨oßer (kleiner), also zeigt der Pfeil nach rechts (links). Die Geschwindigkeit ist umso gr¨oßer, je gr¨oßer der Betrag von f(N) ist. Dies kann man durch verschieden große Pfeile andeuten. Die Bewegung von N(t) folgt dann diesen Pfeilen. Daraus l¨aßt sich qualitativ der Zeitverlauf ablesen, auch wenn man die Differentialgleichung nicht explizit l¨osen kann.
Fixpunkte und Stabilit¨at
Wie im diskreten Fall interessieren wir uns f¨ur Werte, auf die sich die Gr¨oßeN(t) nach langer Zeit einstellt. Offensichtlich ¨andert sich N(t) gerade an den Punkten nicht mehr, an denen die Geschwindigkeitsfunktion f(N) = 0 ist. Geometrisch sind dies die Schnittpunkte von f(N) mit derx-Achse. Die Stabilit¨at der Gleichgewichtspunkte (bzw. Fixpunkte) und andere elementare Eigenschaften lassen sich direkt aus dem Phasenliniendiagramm erschließen:
• Ein GleichgewichtspunktN∗ istlokal stabil(anziehend), wenn die Pfeile in beiden Rich- tungen auf ihn zulaufen. Eine mathematisch hinreichnde Bedingung daf¨ur ist, ist dass die Ableitung f0(N) = dNd f(N) < 0 ist. Ist dagegen f0(N∗) > 0, so laufen die Pfeile nach links und rechts vom Fixpunkt weg und das Gleichgewicht istinstabil (abstoßend).
F¨urf00(N∗) = 0 h¨angt die Stabilit¨at von h¨oheren Ableitungen ab (z.B. ist ein internes Gleichgewicht in diesem Fall instabil, wenn f00(N∗)6= 0, usw.). Generell ist ein Gleich- gewichtN∗ offensichtlich genau dann lokal stabil wenn f¨ur einδ >0 und alle 0< < δ gilt: f(N∗+)<0 undf(N∗−)>0.
• Sei f(N) kontinuierlich differenzierbar. Wir nennen ein instabiles Gleichgewicht einen Bruchpunkt wenn f¨ur kleine f(N∗+) >0 und f(N∗−) < 0 ist. Wenn f(N) nur eine endliche Anzahl von Gleichgewichten hat (genauer: wenn es keine Intervalle mit f(N) = 0 gibt), dann sind asymptotisch stabile Gleichgewichte und Bruchpunkte immer
Arbeitsblatt 2 zur Vorlesung WS 2020/2021 15 alternierend (d.h. es gibt genau einen stabilen Fixpunkt zwischen zwei Bruchpunkten und umgekehrt).
• Wenn alle Fixpunkte entweder asymptotisch stabile Gleichgewichte oder Bruchpunkte sind (der generische Fall), dann erstreckt sich der Anziehungsbereich jedes stabilen Gleichgewichts bis zu den benachbarten Bruchpunkten oder bis zu den Grenzen des Parameterraums.
Man beachte: ˙N(t) ist stets die Ableitung vonN nach der Zeitt, w¨ahrendf0(N) die Ableitung von f nach der dynamischen Gr¨oßeN ist.
Modellbildung III: Erntemodelle
Betrachte eine Fischpopulation mit einer Dynamik gem¨aß logistischem Wachstum. Wir m¨och- ten diese Population m¨oglichst optimal als nat¨urliche Ressource nutzen und sind deshalb an einer Fischfang-Strategie (oder Erntestrategie) interessiert, die uns langfristig einen großen und stabilen Ertrag verspricht. Zwei verschiedene Strategien stehen zu Auswahl:
1. ¨Uber Fischereiquoten wird eine maximale Anzahl H an Fischen festgelegt, die in jedem Jahr gefangen werden d¨urfen (sog.constant rate harvesting). Wenn bis zu dieser Grenze gefischt wird, kann die Dynamik der Population durch folgende Differentialgleichung beschrieben werden:
N˙ =rN 1−N
K
−H (f¨urN >0), N˙ = 0 (f¨urN = 0). (14) 2. Alternativ kann man die Fangmenge von der Gr¨oße des Bestands abh¨angig machen (relative rate harvesting). In der Praxis geschieht das zum Beispiel ¨uber eine Begrenzung der Anzahl und Gr¨oße von Fischerbooten.
N˙ =rN
1−N K
−EN, (15)
wobei E den zul¨assigen Aufwand f¨ur Fischerei beschreibt und EN die resultierende Fangmenge.
Wir definieren den gr¨oßten nachhaltigen Ertrag (maximum sustainable yield, MSY) als den gr¨oßten Ertrag, der dem Bestand dauerhaft entnommen werden kann. Dabei ist einnachhalti- ger Ertragallgemein die Fangmenge an einem stabilen Gleichgewichtspunkt des dynamischen Systems. Der MSY ist dann das Maximum dieser Fangmenge in Abh¨angigkeit der Modellpa- rameterH oder E, die die Erntestrategie bestimmen.
Aufgabe 2.1:
(a) Berechne f¨ur beide Differentialgleichungen die Gleichgewichtspunkte und ihre Stabilit¨at (mit Hilfe eines Phasenliniendiagramms). Welche qualitativ unterschiedlichen dynami- schen Szenarien kann man f¨ur verschiedene Werte von H und E bekommen?
(b) Fertige f¨ur beide Strategien eine Abbildung an, die die Lage aller (stabilen und nicht stabilen) Gleichgewichtspunkte als Funktion vonHbzw.E(= x-Achsen Variable) zeigt.
(c) Was ist der MSY und f¨ur welche Werte vonH und E bekommen wir ihn?
Arbeitsblatt 2 zur Vorlesung WS 2020/2021 16 Modellanalyse III: Bifurkationen
F¨ur eine qualitative Modellanalyse ist es von zentralem Interesse, wie sich die Gleichge- wichtsstruktur ver¨andert, wenn wir bestimmte Modellparameter ¨andern. Wie wir f¨ur die Erntemodelle gesehen haben, k¨onnen Gleichgewichte verschwinden oder neue Gleichgewich- te entstehen, oder Gleichgewichte k¨onnen kollidieren und dabei ihre Stabilit¨at austauschen.
Diese qualitativen ¨Anderungen in der Gleichgewichtsstruktur dynamischer Systeme werden Bifurkationen genannt. Man kann diese “Dynamik der Gleichgewichte” in einem sogenannten Bifurkationsdiagramm darstellen und allgemein verschiedene Typen von Bifurkationen unter- scheiden. F¨ur eindimensionale dynamische Systeme in kontinuierlicher Zeit sind drei Typen von Bifurkationen besonders wichtig:
1. Transkritische Bifurkation (transcritical bifurcation): zwei Gleichgewichte kollidieren (¨uberkreuzen sich) und tauschen ihre Stabilit¨at aus. DieNormalform der transkritischen Bifurkation mit Bifurkationsparameter α im Punktx=α= 0 lautet
˙
x=αx−x2. (16)
Dieses System hat zwei Gleichgewichte bei x∗1 = 0 und x∗2 =α, die f¨urα= 0 im Punkt x= 0 kollidieren. F¨urα <0 ist das Gleichgewichtx∗2 =α stabil und das Gleichgewicht x∗1= 0 instabil; f¨urα >0 ist x∗1 = 0 lokal stabil und x∗2 =α instabil.
2. Sattel-Knoten Bifurkation (saddle-node bifurcation): zwei Gleichgewichte mit unter- schiedlicher Stabilit¨at kollidieren und l¨oschen sich gegenseitig aus – bzw. es entstehen zwei Gleichgewichte aus dem “Nichts”. Die Normalform lautet
˙
x=α−x2. (17)
F¨ur α < 0 gibt es kein Gleichgewicht. Bei α = 0 entstehen im Punkt x = 0 zwei Gleichgewichte, f¨ur α > 0 liegen sie bei x∗1 = −√
α (instabil) und bei x∗2 = √
α (lokal stabil).
3. Heugabelbifurkation (pitchfork bifurcation): drei Gleichgewichte kollidieren. Daf¨ur gibt es zwei M¨oglichkeiten. Bei einer superkritischen Heugabelbifurkation sind die dei Gleich- gewichte stabil – instabil – stabil und werden zu einem einzigen stabilen Gleichgewicht.
Die Normalform lautet
˙
x=αx−x3. (18)
F¨ur die subkritischen Fall sind die drei Gleichgewichte instabil – stabil – instabil und werden zu einem instabilen Gleichgewicht,
˙
x=αx+x3. (19)
4. Im Prinzip sind auch Bifurkationen noch h¨oherer Ordnung m¨oglich, bei denen vier oder mehr Gleichgewichte kollidieren. Sie spielen aber in der Praxis keine Rolle.
5. Außer durch Bifurkationen kann sich die Gleichgewichtsstruktur des dynamischen Sy- stems auch ¨andern, wenn Gleichgewichte in den relevanten biologischen Zustandsraum hineinlaufen oder ihn verlassen. F¨ur die Populationsdichte N ist die relevante unte- re Grenze des Zustandsraums normalerweise N = 0. Auf der anderen Seite k¨onnen Gleichgewichte auch im Unendlichen entstehen oder dorthin verschwinden. Allerdings gilt f¨ur realistische biologische Modelle gew¨ohnlich limN→∞f(N) << 0, sodass dies nicht m¨oglich ist.
Arbeitsblatt 2 zur Vorlesung WS 2020/2021 17 Aufgabe 2.2:
(a) Welche Bifurkationen treten f¨ur absolute und relative rate harvesting auf? Diskutiere auch die Konsequenzen beider Bifurkationstypen, wenn man die Ernterate variiert - ggf. auch ¨uber den Bifurkationspunkt hinaus. Welche Strategie ist zu bef¨urworten?
(b) F¨urrelative rate harvesting bekommt man die maximale Gesamt-Fangmenge f¨ur einen mittleren Wert des gesamten zul¨assigen Fischereiaufwands. Man kann sich aber auch uberlegen, was passiert, wenn mehrere Fischereiunternehmen oder mehrere L¨¨ ander um den selben Bestand konkurrieren. Wie ¨andert sich dann der individuelle Ertrag mit dem individuellen Aufwand eines einzigen Unternehmens (oder Landes)? Was bedeutet das f¨ur die Entwicklung des Gesamtertrags?
Bedeutung der Bifurkationstheorie: Kleine Ursachen k¨onnen große Effekte haben.Dies ist eine typische Eigenschaft von ¨Okosystemen, wo man diese pl¨otzlichen ¨Uberg¨ange h¨aufig beobachtet. Die Tatsache, dass kleine stetige ¨Anderungen große sprunghafte Auswirkungen haben k¨onnen ist f¨ur die meisten Menschen kontraintuitiv. Dies kann gef¨ahrliche Konsequen- zen haben wenn ein ¨Okosystem “umkippt” und beispielsweise Arten aussterben. Einblicke in die Bifurkationsstruktur nat¨urlicher Systeme zu geben, ist deshalb eine der wichtigsten Auf- gaben der theoretischen ¨Okologie. In vielen F¨allen ergibt sich die Existenz eines bestimmten Bifurkationspunkts aus der Struktur der Wechselwirkungen und mak kann robuste qualitati- ve Voraussagen erhalten. Nat¨urlich wird es oft schwierig sein, quantitative Sch¨atzungen f¨ur die genaue Positionen der Bifurkationspunkte zu erhalten. Zudem weisen nat¨urliche Syste- me immer auch ¨okologische und demographische Stochastizit¨at auf. Deshalb ist es ratsam, stets einen gen¨ugend großen Sicherheitsabstand zu einem katastrophalen Bifurkationspunkt einzuhalten.
Ubungsaufgaben 2¨
Aufgabe 2.3: Die rechte Seite einer Differentialgleichung sei durch die Funktion g(x) = sin(πx/2)
gegeben.
(a) Zeichnen Sie ein Phasenliniendiagramm f¨urx zwischen 0 und 4. F¨ur welche Werte von x ¨andert sichx am schnellsten?
(b) Welche Gleichgewichtspunkte gibt es und was k¨onnen Sie ¨uber ihre Stabilit¨at aussagen?
(c) Machen Sie anhand der qualitativen Informationen eine grobe Skizze des Zeitverlaufs f¨ur Startwerte x0 = 0.5 and x0 = 3.5. Gibt es Wendepunkte im Zeitverlauf x(t) und wenn ja, wo liegen sie?
Aufgabe 2.4: Chemostat
Ein Chemostat ist ein Apparat, um Bakterien unter Durchflußbedingungen zu kultivieren.
Dabei wird kontinuierlich frische N¨ahrl¨osung zugef¨uhrt; mit derselben Rate wird Kulturme- dium – mitsamt den darin enthaltenen Bakterien – abgef¨uhrt, so daß das Volumen insgesamt konstant bleibt.
Arbeitsblatt 2 zur Vorlesung WS 2020/2021 18
Wir nehmen an, daß die Bakterien logistisch wachsen mit Parametern λundK, und das Kulturmedium gut ger¨uhrt wird, so daß die Zellen ¨uberall gleichm¨aßig verteilt sind. Seixdie Dichte der Bakterienkultur undα die Auswaschrate (Abflussrate).
(a) Stellen Sie die ODE des Modells auf. Welche Modellannahmen f¨uhren zu dieser Glei- chung?
(b) Zeichnen Sie das Phasenliniendiagramm. F¨ur welchen Wert vonx ist die ¨Anderung von x maximal? F¨ur welche Werte vonx¨andert sichxnicht? Ermitteln Sie – graphisch und rechnerisch – die Stabilit¨at der Gleichgewichtspunkte.
(c) F¨ur welche Wahl des Anfangswertes x0 wird der Zeitverlauf x(t) einen Wendepunkt haben?
Arbeitsblatt zur 3. Vorlesung, WS 2020/2021 19
2 Effekte diskreter Dynamik in einer Dimension
Systeme in kontinuierlicher und diskreter Zeit
Zwischen diskreten dynamischen Systemen und Differentialgleichungen gibt es zahlreiche Ent- sprechungen, die in der folgenden Tabelle zusammengefasst sind.
diskrete Zeit (n) kontinuierliche Zeit (t) dynamisches System diskrete Iteration Differentialgleichung
xn+1 =f(xn) N˙(t) =f(N)
charakterisiert durch Iterationsfunktionf(x) Geschwindigkeitsfunktion f(N) gibt an Populationsgr¨oße in der Zuwachsrate der Population
n¨achsten Generation
graphischer Ansatz Cobwebbing Phasenliniendiagramm
Gleichgewicht f(x) =x f(N) = 0
Schnittpunkt mit Winkelhalbierender x-Achse lokal stabil wenn |f0(x∗)|<1 f0(N∗)<0
Neben diesen Entsprechungen gibt es aber auch wichtige Unterschiede. Insbesondere sehen wir aus dem Phasenliniendiagramm, dass die Konvergenz zu einem Gleichgewicht im kontinuierli- chen Fall immer monoton ist: ein Gleichgewichtspunkt kann im Zeitverlauf nie ¨ubersprungen werden (mathematisch folgt dies aus der Eindeutigkeit der L¨osungen f¨ur Differentialgleichun- gen). Allgemeiner gilt:
• Zu jedem kontinuierlichen Prozess gibt es eine Menge an analogen diskreten Prozessen.
Wie beim ¨Ubergang vom exponentiellen zum geometrischen Wachstum ergeben sich die entsprechenden diskreten Prozesse einfach dadurch, dass man die Populationsgr¨oße nur in (beliebig gew¨ahlten) diskreten Zeitschritten aufzeichnet. Insbesondere die Gleichge- wichtspunkte sind dann identisch.
• Umgekehrt gibt es nicht zu jedem diskreten Prozess einen analogen kontinuierlichen Prozess, der sich wie oben beschrieben durch eine gew¨ohnliche Differentialgleichung darstellen l¨asst. Daf¨ur werden wir im n¨achsten Abschnitt einige konkrete Beispiele ken- nenlernen.
2.1 Nicht-lineare Prozesse in diskreter Zeit
Analog zum logistischen Wachstum in kontinuierlicher Zeit muss das Populationswachstum auch in biologisch realistischen Modellen in diskreter Zeit bei hohen Dichten verlangsamt und schließlich gestoppt werden (wie schon am Beispiel der Vogelpopulation argumentiert). Daf¨ur m¨ussen die mittlere Anzahl der Geburtenbund der Todedpro Individuum und Zeiteinheit von der Populationsgr¨oße abh¨angen. Die Reproduktionsfunktion nimmt dann die allgemeineren Form
f(x) =x(1 +b(x)−d(x)) =xr(x) (20) an, in derb, d und r selbst Funktionen von x sind. Beim geometrischen Wachstum war die Reproduktionsfunktion eine Gerade. Man nennt den zugeh¨origen Prozess deshalb auch einen
Arbeitsblatt zur 3. Vorlesung, WS 2020/2021 20 linearen Prozess. Prozesse mit Iterationsfunktionen, die keine Geraden sind, nennt man ent- sprechendnicht-lineare Prozesse. In diesem Abschnitt werden wir Modelle von nicht-linearen Prozessen vorstellen. Wir wollen aber nicht einfach mit den fertigen Formeln beginnen, son- dern diese aus einem biologischen Szenario ableiten.
Modellbildung IV: Fischpopulation
Betrachte eine Fischpopulation mit Gr¨oßeNtf¨ur den erwachsenen (adulten) Bestand. W¨ahrend der Brutsaison produzieren die Adulten Lt = bNt Larven. Wir nehmen an, dass diese Lar- ven w¨ahrend eines Jahres bis zur Fortpflanzungsreife heranwachsen und dass adulte Fische nur eine Saison ¨uberleben. Dann bilden die ¨uberlebenden Larven die adulte Population der n¨achsten Generation. Wir brauchen also ein Modell f¨ur das ¨Uberleben der Larven ¨uber ein Jahr.
1. Angenommen, die Larven entwickeln sich w¨ahrend dieses Jahres gem¨aß gem¨aß eines (zeitlich kontinuierlichen) logistischen Modells ohne Geburten, d.h,
∂Lt(τ)
∂τ =−m1Lt(τ)−m2L2t(τ), (21) wobeim1 die Sterblichkeit aufgrund von extrinsischen (z.B. abiotischen) Faktoren misst and m2 die dichteabh¨angige Komponente (z.B. aufgrund von Ressourcenkonkurrenz zwischen den Larven oder Jungfischen). Mit Lt(0) =bNtund der Dauer ∆ einer Saison wird
Lt(∆) = bNtm1
(bNtm2+m1) exp[m1∆]−bNtm2
. (22)
Damit erhalten wir das sogenannte Beverton-Holt Modell, Nt+1=Lt(∆) = c1Nt
1 +c2Nt
(23) mit positiven Konstanten
c1=bexp[−m1∆] , c2 =bm2
m1 (1−exp[−m1∆]).
2. Ein alternativer Ansatz geht davon aus, dass die Larven nicht prim¨ar untereinander, sondern mit ihren erwachsenen Artgenossen konkurrieren. Dies trifft insbesondere dann zu, wenn adulte Fische Eier und Larven ihrer eigenen Art fressen (was in der Tat f¨ur viele Arten zutrifft, bei denen die Pr¨adation rein gr¨oßenabh¨angig ist). Wir erhalten dann
∂Lt(τ)
∂τ =−m1Lt(τ)−m2NtLt(τ), (24) und
Lt(∆) =bNtexp[−(m1+m2Nt)∆]. (25) Dies f¨uhrt auf das sogenannteRicker Modell,
Nt+1 =Ntexp[r(1−Nt/K)] (26)
mit Konstanten
r= log[b]−m1∆ , K= log[b]−m1∆ m2∆ .
Arbeitsblatt zur 3. Vorlesung, WS 2020/2021 21 3. Schließlich k¨onnen wir auch annehmen, dass Jungfische (unabh¨angig voneinander) mit Wahrscheinlichkeits¨uberleben und der dichteabh¨angige Wettbewerb ausschließlich un- ter den Erwachsenen stattfindet (z.B. um Brutpl¨atze). Dann erhalten wir eine diskrete Version des logistischen Wachstums
Nt+1 =rNt
1−Nt
K
, (27)
wobei r = sb die Zahl der bis zur n¨achsten Saison ¨uberlegenden Larven pro adulten Fisch angibt.
Die Rekrutierung der neuen Generation mittels einer (strikt) monoton steigende Reprodukti- onsfunktion wie beim Beverton-Holt Modell wird als normal-kompensierend (normal compen- sation) bezeichnet: je mehr Eltern es gibt, desto mehr Nachkommen. Sie erreicht eine Asym- ptote vonNt → ∞ bei Lmax(∆) = c1/c2. Im Gegensatz dazu ist die Reproduktionsfunktion des Ricker-Modells nicht monoton. Dasselbe gilt f¨ur das diskrete logistische Wachstumsmodell.
In beiden Modellen ist die Rekrutierung ¨uberkompensierend (over-compensating). Lmax(∆) durchl¨auft ein lokales Maximum und n¨ahert sich f¨urNt→ ∞dem Wert Null. Die biologische Herleitung der Modelle ¨uber die j¨ahrliche Erneuerung eines Fischbestands ist nur ein Bei- spiel. Alle drei Modelle k¨onnen auf viele verschiedene Arten f¨ur unterschiedliche biologische Szenarien abgeleitet werden. Sie sind Standardmodelle der theoretischen ¨Okologie.
Analyse des Beverton-Holt Modells
Betrachte die Reproduktionsfunktion des Beverton-Holt (oder auch Verhulst) Modells, Nt+1=F(Nt) = c1Nt
1 +c2Nt
. (28)
Reproduktionsfunktion und Populationsentwicklung sind in Abb. 5 gezeigt, im Vergleich zum geometrischen Wachstum (c2= 0).
• Im Gleichgewicht gilt
N(1 +c2N) =c1N und damit
N1∗ = 0, N2∗ = c1−1 c2 . F¨urc1 >1 istN2∗ >0 und biologisch relevant.
• Mit
F0(N) = ∂F(N)
∂N = c1
(1 +c2N)2 ≤c1 =F0(0)
und F0(N2∗) = 1/c1<1 f¨urc1>1, wennN2∗ >0 ist. Wir bekommen deshalb monotone Konvergenz gegen N2∗ f¨urc1 >1 und ansonsten monotone Konvergenz nach N1∗= 0.
Arbeitsblatt zur 3. Vorlesung, WS 2020/2021 22
0 10000 20000 30000 40000 50000 x
2000 4000 6000 8000 10000 12000
fHxL
0 50 100 150 200
Zeit t 0
2000 4000 6000 8000 10000 12000
Population
Abbildung 5: Reproduktionsfunktionen (links) und Populationsentwicklung (rechts) f¨ur das Beverton-Holt Modell mit N0 = 1, c1 = 1.1, c2 = 1/10000 und geometrisches Wachstum mitr =c1 = 1,1. Zeit in Generationen gemessen; jede 5. Generation ist gezeigt (Rauten = Beverton-Holt, Sterne = geometrisch).
Analyse des Ricker Modells
Wir betrachten die Reproduktionsfunktion
Nt+1 =F(Nt) =Ntexp[r(1−Nt/K)]. (29)
• Wir erhalten die Gleichgewichte
N1∗ = 0, N2∗=K .
• Die Ableitung ist
F0(N) =
1−N r K
exp[r(1−N/K)]. und damit
Λ1=F0(0) = exp[r]>1, (30)
Λ2=F0(K) = 1−r. (31)
• Wir haben ein eindeutiges Maximum von F(N) bei Nmax = K/r (und ein Minimum bei Nmin = 0).
Damit k¨onnen wir drei dynamische Regimes f¨ur das Ricker Modell unterscheiden.
1. F¨urr <1 enth¨alt das IntervallI = [0, Nmax] beide Gleichgewichtspunkte. DaF(N) inI monoton steigt erhalten wir monotone Konvergenz zum stabilen GleichgewichtN2∗ =K f¨ur jeden Startwert N0 ∈I. F¨ur StartwerteN0 ∈/ I haben wirF(N0)∈I und dasselbe gilt nach dem ersten Schritt.
2. F¨ur 1< r <2 haben wir keine monotone Konvergenz mehr, aber immer noch |Λ2|<1.
Daher ist N2∗ = K immer noch lokal stabil. Die Populationsgr¨oße n¨ahert sich dem Gleichgewicht in ged¨ampften Schwingungen.
3. F¨ur r > 2 ist |Λ2| > 1 und wir haben kein stabiles Gleichgewicht mehr. Insbesondere erhalten wir f¨ur das Gleichgewicht beiN2∗ =K oszillierende Divergenz. Da das Ricker- Modell eine endliche maximale Populationsgr¨oße hat, stellt sich die Frage, wohin sich der Zeitverlauf entwickelt.
Arbeitsblatt zur 3. Vorlesung, WS 2020/2021 23 Ubungsaufgaben 3¨
Aufgabe 3.1: Denken Sie sich eine Bakterienkultur, die mit r = 1.5 geometrisch w¨achst.
In dieser Kultur tritt eine Mutante auf, die mit r = 2.0 geometrisch w¨achst; die Mutante w¨achst also schneller als der Wildtyp. Wie sieht die Reproduktionsfunktion f¨ur den Anteil der Mutanten an der Gesamtpopulation aus? (Hinweise: Man bezeichne mitwndie Gr¨oße der Wildtyppopulation (in Generation n), mit mn die Gr¨oße der Mutantenpopulation und mit Nn:=wn+mn die Gr¨oße der Gesamtpopulation. Betrachten Sie nun den Anteilpn der Mu- tanten an der Gesamtpopulation. Versuchen Sie nun,pn+1 als Funktion vonpnauszudr¨ucken.
Verwenden Sie dazu das geometrische Wachstumsgesetz f¨urwn und mn sowie die Tatsache, daß der Anteil der Wildtypen gleich 1−pn ist.) Kommt Ihnen die Gleichung bekannt vor?
Aufgabe 3.2: (Biologische Sch¨adlingsbek¨ampfung) Die Generationen eines sch¨adlichen Insektes sind streng getrennt. Jede Generation besteht aus gleich vielen M¨annchen und Weib- chen. Weibchen und M¨annchen paaren sich rein zuf¨allig und nur ein einziges Mal. Seimndie Zahl der fruchtbaren M¨annchen in der n−ten Generation, 2r (r > 1) sei die mittlere Zahl an Nachkommen pro (fruchtbarer) Paarung. Pro Generation werden s >0 sterile M¨annchen freigelassen, die sich ansonsten wie die anderen M¨annchen verhalten. Ziel der Aktion ist es, die Insektenpopulation aussterben zu lassen.
(a) Stellen Sie die Reproduktionsfunktion f¨ur mn auf. (Hinweise: Da sich jedes Weibchen zuf¨allig mit irgendeinem M¨annchen der Population paart, ist die Zahl der fruchtbaren Paarungen gleich der Zahl der Weibchen mal dem Anteil der fruchtbaren M¨annchen an den M¨annchen insgesamt; die Gesamtzahl der M¨annchen istmn+s.)
(b) Berechnen Sie die Fixpunkte der Reproduktionsfunktion und ihre Stabilit¨at. Zeichnen Sie die Reproduktionsfunktion f¨ur das Beispiel r = 3/2, s = 1/2 und lesen Sie die Stabilit¨at der Fixpunkte aus der Zeichnung ab.
(c) Wie groß mußs– f¨ur ein gegebenesm0– gew¨ahlt werden, damit die Insektenpopulation ausstirbt?
Arbeitsblatt zur 4. Vorlesung, WS 2020/2021 24 2.2 Grenzzyklen und Chaos
Modellanalyse IV: Grenzzyklen
Die Grundidee, um weitere Erkenntnisse ¨uber die Langzeitdynamik des Ricker-Modells und
¨ahnlicher Systeme zu gewinnen, ist die Betrachtung der iterierten Abbildung ¨uber k Zeit- schritte,
F(2)(N) =F(F(N)) resp. F(k)(N) =F(. . . F(F(N))) k-fach. (32) Offensichtlich handelt es sich wieder um ein diskretes dynamisches System, das auf die gleiche Weise untersucht werden kann wie das urspr¨ungliche. Hinsichtlich seines Langzeitverhaltens beobachten wir die folgenden elementaren Eigenschaften:
1. Gleichgewichte von F sind auch Gleichgewichte von F(2) und jeder h¨oheren Iteration F(k).
2. F¨ur die Ableitung vonF(2) an den Fixpunkten folgt mit der Kettenregel,
∂F(2)(N)
∂N = ∂F(Z)
∂Z
Z=F(N)·∂F(N)
∂N . (33)
und genauso f¨ur die iterierte Abbildung F(k). Das bedeutet f¨ur die Gleichgewichte N∗
∂F(k)(N)
∂N
N=N∗ =
∂F(N)
∂N N=N∗
k
. (34)
Stabile Gleichgewichte von F bleiben also auch f¨ur h¨ohere Iterationen stabil und insta- bile Gleichgewichte bleiben instabil.
3. Weitere (stabile oder instabile) Gleichgewichte von iterierten AbbildungenF(k) k¨onnen auftreten, wenn die urspr¨ungliche AbbildungF Zyklen hat.
Definition und grundlegende Eigenschaften: Grenzzyklen
1. Ein Punkt N0 wird Punkt mit Periode k oder k-Zyklus Punkt genannt wenn er ein Gleichgewichtspunkt der k-fach iterierten Abbildung F(k)(N) ist, aber kein Fixpunkt einer Abbildung F(k0) mit 1≤k0 < k. Sein sogenannterOrbit
{N0, F(N0), . . . , F(k−1)(N0)}=:{N0, N1, . . . , Nk−1}
wird der entsprechende k-Zyklus genannt. Alle Punkte Ni in diesem Zyklus sind Fix- punkte von F(k).
2. Ein Grenzzyklus ist lokal stabil genau dann, wenn die zugeh¨origen Zyklus-Punkte lokal stabile Gleichgewichte derk-fach iterierten Abbildung sind. Mit der Kettenregel erhalten wir f¨ur die AbleitungF(k) an allen Zyklus-Punkten
Λ(k) = ∂F(k)
∂N =
k−1
Y
i=0
∂F(N)
∂N N=Ni
. (35)
Der Zyklus ist mithin lokal stabil wenn |Λ(k)|<1 ist und instabil f¨ur|Λ(k)|>1.
Arbeitsblatt zur 4. Vorlesung, WS 2020/2021 25 Beispiel: diskretes logistisches Wachstum
Wir betrachten die Iterationsfunktion f¨ur das diskrete logistische Wachstum Nt+1 =F(Nt) =rNt
1−Nt
K
, (36)
die sich ¨ahnlich zur Ricker Gleichung verh¨alt, aber leichter zu analysieren ist. F(N) hat ein Maximum bei N = K/2 mit F(K/2) = rK/4. Es gibt zwei Gleichgewichte N1∗ = 0 und N2∗ =K(r−1)/r. Mit F0(N) =r(1−2N/K) erhalten wir F0(0) =r und F0(N2∗) = 2−r.
1. F¨ur 0 < r <1 ist N1∗ = 0 das einzige stabile Gleichgewicht und die Population stirbt aus.
2. F¨ur 1 < r <3 entsteht ein zweites, stabiles Gleichgewicht N2∗ w¨ahrend N1∗ instabil ist (transkritische Bifurkation im Punkt r = 1). Die Konvergenz nach N2∗ ist f¨ur r < 2 monoton und f¨urr >2 oszillierend.
3. F¨urr >3 sind beide GleichgewichteN1∗ undN2∗ instabil und wir erwarten Grenzzyklen oder noch komplexeres Langzeit-Verhalten.
F¨ur ein wohldefiniertes biologisches Modell muss r ≤ 4 sein, da sonst F(N) > K werden kann, was negative Populationsgr¨oßen in der n¨achsten Generation zur Folge h¨atte. Um die Dynamik im Detail zu studieren, betrachten wir den Fall K= 1. Damit wird:
F(N) =rN(1−N) (37)
F(2)(N) =r(rN(1−N))(1−rN(1−N)) =r2N(1−N)(1−rN +rN2) (38)
∂F(2)(N)
∂N =r 1−2rN(1−N)
r(1−2N) (39)
(Wir haben allgemein die SymmetrieF(k)(1−N) =F(k)(N).) Wir bekommen Gleichgewichte F(2)(N) =N f¨ur
N1∗= 0 , N2∗ = r−1
r , N3,4∗ = 1 +r±p
(r−1)2−4
2r . (40)
Offensichtlich existierenN3,4∗ f¨urr≥3. Weiter giltF(N3∗) =N4∗ und umgekehrtF(N4∗) =N3∗. Die Ableitung ist
Λ(2)1 =r2 Λ(2)2 = (2−r)2 , Λ(2)3,4 = 4 + 2r−r2 (41)
• Bei r= 3 istN2∗ =N3∗ =N4∗. Wenn wir rweiter erh¨ohen, ¨andert das GleichgewichtN2∗ seine Stabilit¨at (stabil → instabil). Gleichzeitig entstehen f¨ur die iterierte Abbildung F(2)(N) zwei neue Gleichgewichte, N3∗ und N4∗. Beide sind stabil: |Λ(2)3,4| < 1 f¨ur 3 <
r < 1 +√
6 ≈3.45. Das ist die typische Signatur einer Heugabel-Bifurkation. F¨ur die urspr¨ungliche AbbildungF(N) ¨andert sich die Zahl der Gleichgewichte nicht. Die neuen Gleichgewichte von F(2) bilden f¨urF(N) einen stabilen Grenzzyklus mit Periode 2.
• F¨ur r > 1 +√
6 ≈ 3.45 wird Λ(2)3,4 <−1 und die Gleichgewichte N3∗ und N4∗ von F(2) werden instabil. F¨ur die iterierte Abbildung F(4) bedeutet das, dass die Ableitung an