auf humane T-Zellen Einfluss von Vitamin D und Vitamin A
154
0
0
Volltext
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(19)
(20)
(21)
(22)
(23)
(25)
(26)
(27)
(28)
(29)
(30)
(32)
(33)
(34)
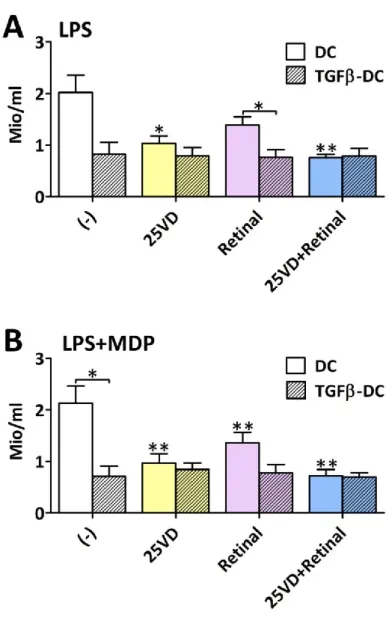
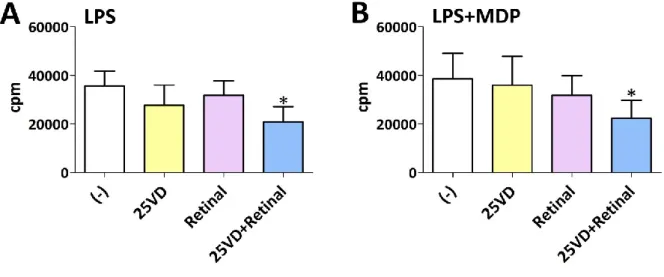
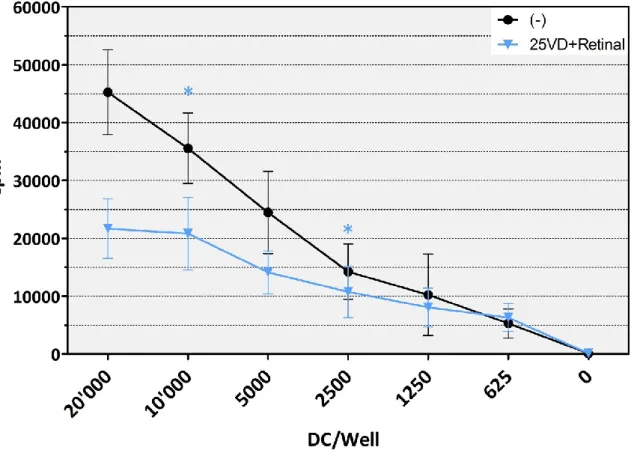
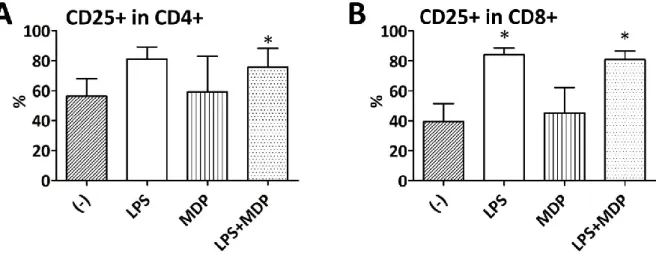
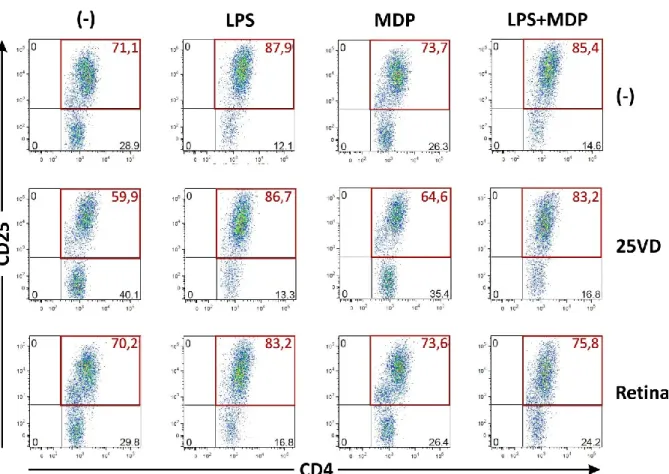
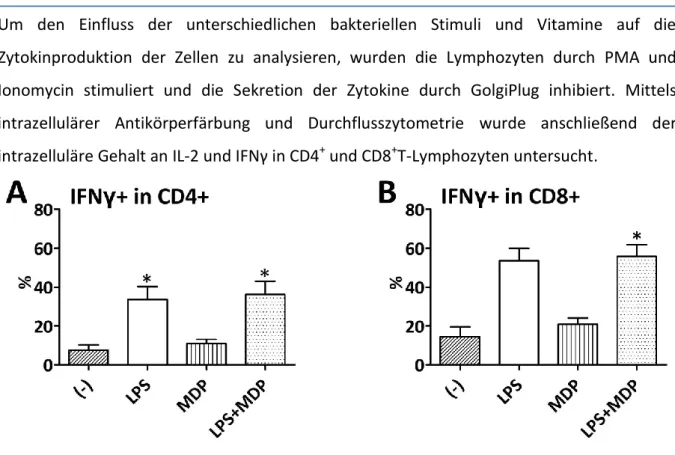
Abbildung
+7
ÄHNLICHE DOKUMENTE
Vitamin D spielt eine zentrale Rolle für die Knochengesund- heit und damit für die Osteoporose-Prophylaxe.. Auch das Immunsystem profitiert von ausreichend Vitamin D, denn es
Zwischen Ende Oktober und März kann unser Körper selbst nicht ausreichend Vitamin D
Dies sollte bei der Vit D Dosisanpassung im Falle einer Unterversorgung/ Mangel berücksichtig werden, damit es zu keiner unbeabsichtigten Überdosierung kommt.. Klinisch bewährt
• Über die Ernährung oder Sonnenexposition ist eine Überdosierung nicht möglich. Multivitaminbrausetabletten) oder hochdosierten Vitamin- D-Supplementen (Tropfen); insbesondere
Laif ® 900 Balance darf nicht eingenommen werden bei bekannter Allergie gegenüber Johanniskraut oder einem der sonstigen Bestandteile?. Hinweise: Ausreichende Erfahrungen über
Eine aktuelle Untersuchung der Universität von Tel Aviv zeigt, dass bei Asthmapatienten mit einem Vi- tamin-D-Dezifit das Risiko, einen Asthmaanfall zu er- leiden, um 25 Prozent
Vielfältige Wirkungen – Es ist eine Folge der geringen Sonnenexposition: Ausgerechnet mit dem Vitamin, das streng genommen gar keines ist, weil der Körper es selbst
Entscheidend für diese Änderungen waren – wie die DGE mitteilte – Studien zur Knochengesundheit, die zeig- ten, dass das Risiko für Kno- chenbrüche und Stürze bei Senioren durch
