Die Aldo-Keto-Reduktase AKR1B7: Koexpression mit Renin in der Mausniere und Einfluss auf die Reninexpression und -sekretion
118
0
0
Volltext
(2)
(3)
(4)
(5)
(6)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(18)
(19)
(20)
(21)
(22)
(23)
(24)
(25)
(26)
(27)
(28)
(29)
(30)
(31)
(32)
(33)
(34)
(35)
(36)
(37)
(38)
Abbildung

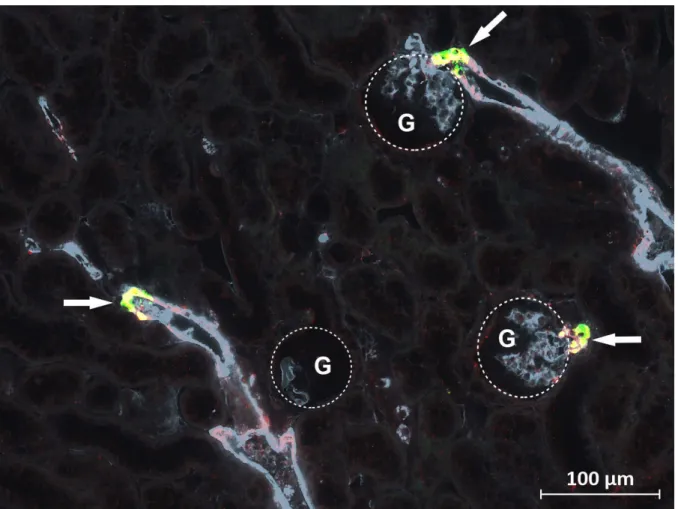
![Abbildung 3.1: [Gesamtansicht] 3D-Rekonstruktion der Glattmuskelaktin- (rosa), Renin- (grün) und AKR1B7- (rot) Immunreaktivität im repräsentativen Gefäßbaumabschnitt einer Wildtyp-Maus [NS] mit](https://thumb-eu.123doks.com/thumbv2/1library_info/3945369.1534171/50.892.180.717.601.1097/abbildung-gesamtansicht-rekonstruktion-glattmuskelaktin-immunreaktivität-repräsentativen-gefäßbaumabschnitt-wildtyp.webp)
+7
![Abbildung 3.2: [Gesamtansicht] 3D-Rekonstruktion der Glattmuskelaktin- (rosa) Immunreaktivität im repräsentativen Gefäßbaumabschnitt einer Wildtyp-Maus [NS] mit Darstellung der Glomeruli und der Nierenkapsel (gelb)](https://thumb-eu.123doks.com/thumbv2/1library_info/3945369.1534171/51.892.183.719.79.1086/gesamtansicht-rekonstruktion-glattmuskelaktin-immunreaktivität-repräsentativen-gefäßbaumabschnitt-darstellung-nierenkapsel.webp)
![Abbildung 3.5: [Detailansicht] Immunfluoreszenzfärbung am Paraffinschnitt einer Wildtyp-Maus [NS]](https://thumb-eu.123doks.com/thumbv2/1library_info/3945369.1534171/54.892.94.805.359.929/abbildung-detailansicht-immunfluoreszenzfärbung-paraffinschnitt-wildtyp-maus-ns.webp)
![Abbildung 3.7: [Detailansicht] Immunfluoreszenzfärbung am Paraffinschnitt einer Wildtyp-Maus [HS]](https://thumb-eu.123doks.com/thumbv2/1library_info/3945369.1534171/56.892.93.807.234.802/abbildung-detailansicht-immunfluoreszenzfärbung-paraffinschnitt-wildtyp-maus-hs.webp)
![Abbildung 3.13: [Detailansicht] 3D- Rekonstruktion der Glattmuskelaktin- (rosa) Immunreaktivität mehrerer afferenter Arteriolen einer Aldosteronsynthase-Knockout-Maus mit Glomeruli (gelb) und einer efferenten Arteriole](https://thumb-eu.123doks.com/thumbv2/1library_info/3945369.1534171/62.892.126.755.316.963/abbildung-detailansicht-rekonstruktion-glattmuskelaktin-immunreaktivität-afferenter-arteriolen-aldosteronsynthase.webp)
![Abbildung 3.14: [Detailansicht] 3D- Rekonstruktion der Glattmuskelaktin- (rosa) Immunreaktivität mehrerer afferenter Arteriolen einer Aldosteronsynthase-Knockout-Maus mit Glomeruli (gelb) und einer efferenten Arteriole](https://thumb-eu.123doks.com/thumbv2/1library_info/3945369.1534171/63.892.134.762.342.994/abbildung-detailansicht-rekonstruktion-glattmuskelaktin-immunreaktivität-afferenter-arteriolen-aldosteronsynthase.webp)
ÄHNLICHE DOKUMENTE
• Prolaktinkonzentrationen im Plasma, die 150 – 200 ng/mL überschreiten weisen auf Prolaktinom als Ursache der Hyperprolaktinämie hin. • Prolaktinkonzentrationen im Plasma,
• Latente Formen der Hyperprolaktinämie, die man im Basalwert allein nicht erfassen kann, die jedoch durchaus negative Auswirkungen auf die Follikel- reifung haben, diagnostiziert
Dies führte zu der Hypothese, dass eine ß 2 -adrenerge Stimulation, auch durch diese Medikamente, über eine Aktivierung des RAS zu einer vermehrten EPO-Produktion führt... Auf
a quantitative polymerase chain reaction had to be developed to measure renin mRNA. Juxtaglomerular cells were isolated and cultured for 2 days under various conditions, and renin
262 (Renal Fluid Electrolyte Physiol. 31): F397-F402, 1992.- To find out whether calmodulin activity could be a common denominator for the cellular control of renin
In our own studies we have found that T P A enhances transmembrane calcium influx into isolated juxtaglomerular cells (Kurtz et al. Despite the uncertainty about the way
[r]
[r]
