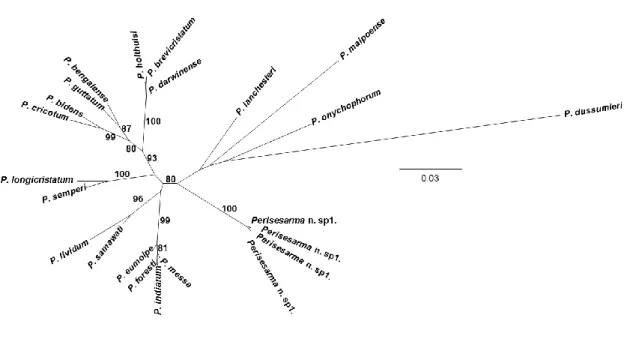
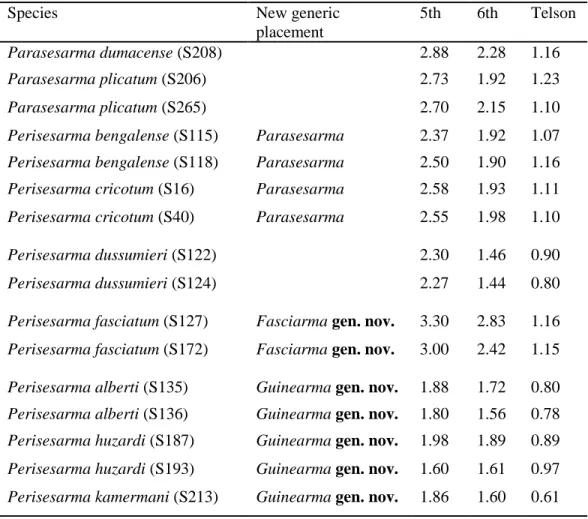
Molecular phylogeny, phylogeography and taxonomic revision of species of the genus Perisesarma De Man, 1895 (Crustacea: Decapoda: Brachyura: Sesarmidae)
206
0
0
Volltext
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(18)
(19)
(20)
(21)
(22)
(23)
(24)
(25)
(26)
(27)
(28)
(29)
(30)
(31)
(32)
(33)
(34)
(35)
(36)
(37)
(38)
(39)
(40)
(41)
(42)
(43)
(44)
Abbildung



+7





ÄHNLICHE DOKUMENTE
Sticherus pteridellus is characterized by having the 1 st axes as long as to slightly longer than the 2 nd , thin rhizomes with scattered, ovate, stoutly hyaline scales,
Legs mostly dark yellow ex- cept tarsus blackish brown; anterior face of femora and tibiae covered in black scales, posterior face of femora and tibiae covered in white
[r]
Redescription. Male: Body length 3 mm. Head: Eye bare. Frontal vitta brownish, reddish on anterior margin. Interocular space 0.4 mm. Two pairs of fr, with two in- tercalated
Thorax: Pronotum side fully punctate; anterior angles slightly acute; marginal groove on anterior margin occu- pying 1/3 of pronotum anterior border; median groove
Eyes dorsolaterally situated on head; eye diameter usually be- tween one-fourth and one-third the length of lateral ce- phalic margin in full-face view, rarely more; scape sur-
as long as wide; antennomere 9 moderately projecting apically, 1.3 × as long as wide; all elytral series weakly but sharply impressed; trochanters with- out tufts of yellowish
The male imago can be separated from other members of the genus by having palpomere 2 very long, its length more than half the length of palpomere 3 and sensorial pit with
