Modellierung des Wachstums und der Ertragsbildung in komplexen Agroökosy- stemmodellen, dargestellt am Beispiel Winterweizen und Winterroggen
Wilfried Mirschel, Alfred Schultz und Jürgen Pommerening, Müncheberg
Zusammenfassung
Ausgehend von prinzipiellen Überlegungen zur Einordnung von Agroökosystemmodellen in eine Hierarchie von Modellen für Agrarflächen wird die zentrale Stellung von Kultur- pflanzenmodellen herausgearbeitet. Anschließend wird der derzeitige Entwicklungsstand des Wachstums- und Ertragsbildungsmodells TRITSIM für Winterweizen vorgestellt. Dabei werden Simulationsergebnisse für die Brandenburger Standorte Müncheberg und Hohenfi- now der Jahre 1987 bis 1992 präsentiert und verglichen. Es werden Möglichkeiten der Übertragbarkeit von Modellstrukturen des Winterweizens auf andere Wintergetreidearten diskutiert und erste Ergebnisse der Modellierung des Winterroggens vorgestellt. Ab- schließend werden mögliche Modellvereinfachungen vorgeschlagen und deren Aus- wirkungen auf die Vorhersagegenauigkeit ausgewählter Pflanzenzustandsvariablen des Winterweizens analysiert.
Summary
Starting from some principal reflections regarding the classification of agroecosystem models within a hierarchy of models for agricultural areas the central position of crop models is emphazised. After that the actual development state of the TRITSIM crop model is introduced. Simulation results are presented and compared for the sites of Hohenfinow and Müncheberg for several years between 1987 and 1992. Possibilities to transfer the TRITSIM model structure and processes to other cereals will be discussed and some preliminary results for winter rye presented. Finally possible changes of the TRITSIM model to reduce the necessary amount of driving forces and parameters are suggested and the effects on model variables are analysed.
1. Einleitung und Zielstellung
Aufgrund der vielfältigen anthropogenen Einflußnahmen ist in der heutigen Zeit eine Ökosystemare Sicht auf unsere natürliche Umwelt mit allen ihren Teilen (Boden, Atmo- sphäre Wald, Gewässer, Agrarlandschaft, ...) zwingend notwendig. Nur eine komplexe Betrachtung und Berücksichtigung der biologischen und ökologischen Gesetzmäßigkeiten und Wechselwirkungen kann sichern, daß bei anthropogenen Eingriffen in die Natur keine irreversiblen, globalen Fehler gemacht werden.
Modelle können wegen ihres wissensintegrierenden Charakters ein Hilfsmittel sein, Ökosysteme in ihrer Komplexität und Kompliziertheit zu beschreiben, besser zu verstehen sowie die Konsequenzen verschiedener Einflußnahmen auf sie abzuschätzen und zu bewerten. Das trifft auch für den flächenmäßig bedeutsamen, agrarisch genutzten Teil der Landschaft zu.
Die Modellierung von Entwicklung, Wachstum und Ertragsbildung der landwirtschaftlichen Kulturen nimmt bei der Abbildung der stofflichen, energetischen und informatorischen Flüsse auf agrarisch genutzten Flächen eine zentrale Rolle ein.
Bei der Erarbeitung von komplexen Agroökosystemmodellen, mit deren Hilfe diese Flüsse widergespiegelt werden sollen, ist das Modellwahlproblem eines der grundlegenden Probleme. Gut wäre es sicherlich, Modelle zu haben, die mit möglichst wenigen, gut interpretierbaren Parametern auskommen, aber gleichzeitig dynamisch genug sind, um auf Klima-, Umwelt- und Bewirtschaftungsänderungen zu reagieren.
Da es sehr unwahrscheinlich ist, ein derart kompliziertes biologisch-ökologisches System, wie es ein Agroökosystem darstellt, auf einfache Weise beschreiben zu können, gilt es, einen Kompromiß zwischen Vorhersagefähigkeit, erklärendem Wert sowie Parameter- und Eingangsgrößenbedarf der Modelle in Abhängigkeit von Zielstellung und räumlichem Größenbezug der Modelle zu finden.
Die vorliegende Arbeit
• gibt einige prinzipielle Überlegungen zur Einordnung von Agroökosystemmodellen in eine mögliche Hierarchie von Modellen für Agrarflächen wieder,
• stellt den aktuellen Entwicklungsstand des Wachstums- und Ertragsbildungsmodells TRITSIM für Winterweizen vor,
• diskutiert Möglichkeiten der Übertragbarkeit der TRITSIM-Modelistrukturen auf andere Fruchtarten und
• untersucht Möglichkeiten der Modellvereinfachung.
Als Beispiele für die Übertragbarkeit und die Modellvereinfachung dienen ein Kuitur- pflanzenmodell Winterroggen bzw. ein Einschicht-Bodenmodell.
2. Agroökosystemmodelle und ihre Einordnung
Um die Stellung der Agroökosystemmodellierung In der Hierarchie des räumlichen Bezuges von Modellen für Agrarflächen zu fixieren und Mißverständnissen vorzubeugen, sei eine Definition für den Begriff des Agroökosystemmodells vorangestellt.
Unter einem Agroökosystemmodeli verstehen wir - in Anlehnung an die Agroökosy- stemdefinition von Leser (1991) - die formale, mathematisch-kybernetische Abbildung wichtiger Teile der Funktionalität eines real vorhandenen Ausschnittes der Geobiosphäre, der durch ein anthropogen gesteuertes Wirkungsgefüge aus abiotischen und biotischen Faktoren charakterisiert ist, und in dem die stofflichen und energetischen Prozesse einheitlich verlaufen (abgrenzbare anthropogene agrarökologische Grundeinheit).
Das Agroökosystemmodeli muß demnach die zielstellungsrelevanten systembeschrei- benden und dynamisch ablaufenden Prozesse im Wechselwirkungsgefüge Boden - Pflanze - Atmoshpäre - Landnutzung abbilden.
Wie nun ist die Agroökosystemmodellierung in der flächenbezogenen Hierarchie zwischen Punkt und Landschaft einzuordnen? Bei welcher Flächen- und damit Systemgröße sprechen wir von Agroökosystemmodellierung?
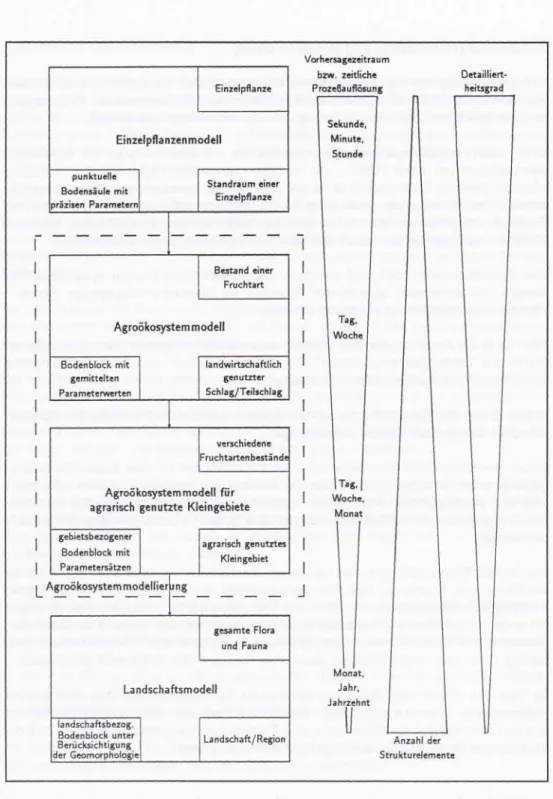
In Abb. 1 wird eine Einordnung der Agroökosystemmodelle in die Hierarchie von Modellen für agrarisch genutzte Flächen vorgeschlagen.
In der Modellhierarchie wird dabei von einem Punktmodell für eine Einzelpflanze aus- gegangen. Im Vordergrund steht hier die Abbildung von morphologischen und inner- pflanzlich-physiologischen Prozessen in Wechselwirkung mit der Umwelt, ohne daß Stoff- und Energiekreisläufe im Wechselwirkungsdreieck Boden - Pflanze - Atmosphäre abgebil- det werden.
Die nächste Ebene wäre dann das Agroökosystemmodell im engeren Sinn. Hier steht die Abbildung von Prozessen des Pflanzenbestandes auf Schlägen bzw. Teilschiägen einschließlich der Abbildung von Stoff- und Energiekreisläufen sowie Wechselwirkungen mit anderen biotischen Komponenten des Agroökosystems (z.B. tierische Schaderreger, Pflanzenkrankheiten, Wildkräuter) im Vordergrund. Die Stoff- und Energiekreisläufe sind, bedingt durch den eingeschränkten räumlichen Bezug, in der Regel nicht geschlossen.
Es folgt die Ebene des Agroökosystemmodells für agrarisch genutzte Kieingebiete (hydrologische Kleineinzugsgebiete). Der Unterschied zum Agroökosystemmodeil im engeren Sinn besteht vor allem darin, daß Fruchtfolgen betrachtet werden und daß die Wasser- und Stoffkreisläufe weitestgehend bilanzierbar sind.
Einzelpflanze
Einzelpflanzenmodell
punktuelle Bodensäule mit präzisen Parametern
Standraum einer Einzelpflanze
Vorhersagezeitraum bzw. zeitliche Prozeßauflösung
Bestand einer Fruchtart
Agroökosystem modeil
Bodenblock mit gemittelten Parameterwerten
landwirtschaftlich genutzter Schlag/Teilschlag
verschiedene Fruchtartenbestände
Agroökosystemmodell für agrarisch genutzte Kleingebiete
gebietsbezogener Boden block mit Parametersätzen
agrarisch genutztes Kleingebiet
i Agroökosystem modellier jng i
gesamte Flora und Fauna
Landschaftsmodell
landschaftsbezog.
Bodenblock unter Berücksichtigung der Geomorphologie
Landschaft/Region
Detaiiliert- heitsgrad
Anzahl der Strukturelemente
Abb. 1: Einordnung der Agroökosystemmodellierung in die Hierarchie des räumlichen Bezuges von Modellen für Agrarflächen
Die abschließende Bezugsebene wird vom Landschaftsmodell gebildet. Dabei wird ein Gebiet bzw. eine ganze Landschaft einschließlich der naturräumlichen Gegebenheiten, Flora und Fauna abgebildet, d.h. sowohl statische als auch dynamische Landschaftsteile.
Hier sind neben den agrarisch genutzten Flächen auch urbane, forstliche und andere Flächen eingeschlossen. Zum Begriff des Landschaftsmodells äußern sich ausführlich
Lutzeetal. (1993).Damit ist der Platz der Agroökosystemmodellierung zwischen dem Einzelpflanzenmodell und dem Landschaftsmodell zu suchen. Sie beinhaltet sowohl Agroökosystemmodelle im engeren Sinn als auch Agroökosystemmodelle für agrarisch genutzte Kleingebiete. In Abhängigkeit von den oben eingeführten Hierarchieebenen kommen mathematische Modelle unterschiedlichen Komplexitätsgrades zum Einsatz. Während es beim Einzel- pflanzenmodell noch möglich ist, detaillierte innerpflanzliche Prozesse mit einem relativ komplizierten mathematischen Gleichungssystem in hoher zeitlicher Auflösung abzubil- den, ist dies auf der Ebene einer Landschaft, bedingt durch fehlende experimentelle Möglichkeiten und numerische Beschränkungen, nicht mehr möglich. Auf der Ebene der Landschaft wird sich die Abbildung aufgrund von begrenzt zur Verfügung stehenden Inputdaten auf einfachere mathematische Beschreibungen beschränken müssen (z. B. auf Wasser-Ertrags-Beziehungen, Temperatursummen-Ontogenese-Beziehungen,...).
3. Wachstums- und Ertragsbildungsmodell TRITSIM für Winterweizen
Das Wachstums- und Ertragsbildungsmodell TRITSIM (TRiTicum SIMulation) wurde in der ersten Hälfte der 80er Jahre vom Forschungszentrum für Bodenfruchtbarkeit Müncheberg in Zusammenarbeit mit dem Zentralinstitut für Kybernetik und Informationsprozesse Berlin entwickelt. Eine erste zusammenfassende Beschreibung ist bei Matthäus etal. (1986) zu finden.
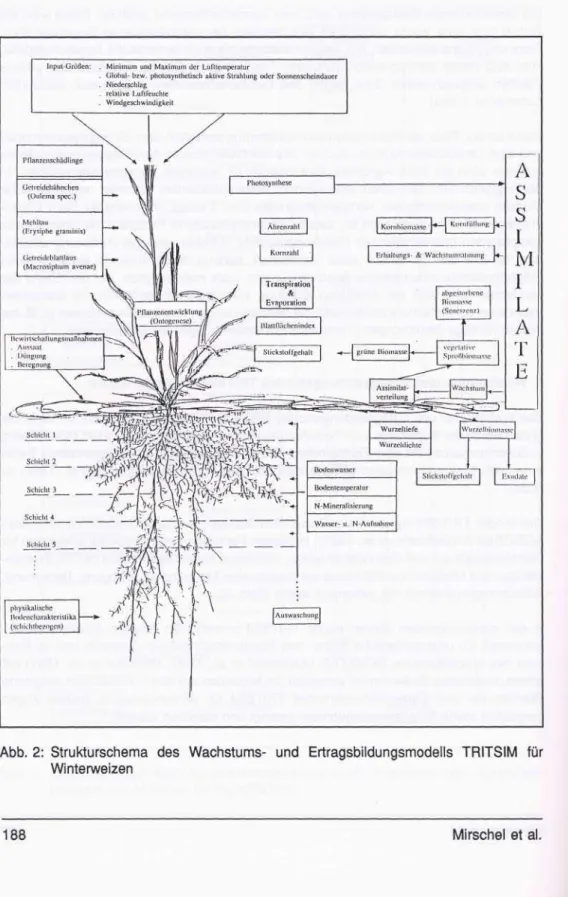
Das Modell TRITSIM war zentraler Bestandteil des Agroökosystemmodells Winterweizen AGROSIM-W (Bellmann etal., 1986), in dessen Rahmen es mit Populationsmodellen für Getreideblattläuse und Getreidehähnchen, Nützlinge, einem Epidemiemodell für Weizen- mehltau und Modellen zur Abbildung agrotechnischer Maßnahmen (Düngung, Beregnung, Schaderregerbekämpfung) gekoppelt wurde (Abb. 2).
In den darauffolgenden Jahren wurde TRITSIM in mehreren Etappen inhaltlich weiter-
entwickelt, für unterschiedliche Klima- und Standortbedingungen überprüft und im Rah-
men des Modellsystems DEMETER (Kartschall et a/., 1990; Matthäus et a/., 1991) mit
einem detaillierten Bodenmodell verknüpft. Im folgenden soll das in DEMETER integrierte
Wachstums- und Ertragsbildungsmodell TRITSIM für Winterweizen in groben Zügen
vorgestellt sowie Simulationsergebnisse gezeigt und diskutiert werden.
Inpui-Größen: . Minimum und Maximum der Lufttemperatur
. Global- bzw. photosynthetisch aktive Strahlung oder Sonnenscheindauer . Niederschlag
. relative Luftfeuchte . Windgeschwindigkeit
^-
IK,,,«™^
1 Erhaltungs- & Wa
-* — 1 grüne Biomasse U
Assimilat- ^^^
— -_^_ Verteilung
J Konifiillnng U~
:hstumsatmung U-
abgestorbene Biomasse (Sencs/cn/.)
SproßWomasse
^^.
A
s
1
IA/l
i
L A r r
B
Abb. 2: Strukturschema des Wachstums- und Ertragsbildungsmodells TRITSIM für Winterweizen
3.1 Modellbeschreibung
Das Modell TRITSIM beschreibt in Tagesschritten vom Auflaufen bis zur Ernte die Entwicklung (Ontogenese), das Wachstum und die Ertragsbildung eines Winterweizen- bestandes unter Freilandbedingungen. Alle Massebilanzierungen werden über den Kohlenstoff vorgenommen. Der Bezug zur Trockenmasse ist über organabhängige Kohlenstoff-Biomasse-Äquivalente möglich. Abb. 2 gibt einen Überblick über die be- rücksichtigten Teilprozesse.
Bei den Teilprozessen nimmt die Ontogenese eine zentrale Stellung ein, da sie die einzelnen Prozesse ein- bzw. ausschaltet, beschleunigt oder abbremst. Die Bedeutung der Ontogenese wird auch aus Tab. 1 deutlich.
Ein weiteres Teilmodell mit zentraler Stellung ist das der Photosynthese als Quelle der täglichen Assimilatbildung. Dabei wird von einer auf die grüne Biomasse bezogenen Photosyntheserate ausgegangen, im Gegensatz zum aus der Literatur bekannten übli- chen LAI-Bezug (z.B. Weir et al., 1984). Die auf die grüne Biomasse bezogene Photo- syntheserate wird durch normierte Strahlungs- und Temperaturcharakteristika (Matthäus et al., 1984), eine Biomasserückkopplungsfunktion, die Tageslänge sowie durch relative Faktoren des Wasser- und Stickstoffversorgungszustandes korrigiert.
Die täglich gebildeten Assimilate werden modellintern in einen Assimilatpool eingespei- chert. Dieser wird täglich bis zu einer von der grünen Biomasse abhängigen unteren Ausschöpfungsschwelle durch die assimilatverbrauchenden Prozesse entleert. Die Entleerung erfolgt hierarchisch in der Reihenfolge Erhaltungsatmung, Kornfüllung, Wachstumsatmung und Wachstum. Die um die Wachstumsrate vermehrte vegetative Biomasse wird über ein ontogeneseabhängiges Verhältnis in vegetative Sproß- und Wurzelbiomasse aufgeteilt. Neben dem Wurzelzuwachs werden eine Wurzelsterberate und eine Rate für Exsudatausscheidungen berücksichtigt. Die Exsudate bilden für das Gesamtsystem einen echten Masseverlust.
In Abhängigkeit vom Alterungsprozeß wird die vegetative Sproßbiomasse in grüne, damit assimilierende, und abgestorbene, photosynthetisch nicht mehr aktive Biomasse aufge- spalten.
Die grüne Biomasse ist Grundlage für die modellmäßige Berechnung sowohl des Blatt- flächenindex als auch der Ährenzahl. Letztere ergibt sich aus der Dynamik des Bio- massezuwachses zwischen den Entwicklungsstadien Schossen (DC 31) und Blüte (DC 61). Aus Ährenzahl und sortenabhängiger Kornzahl pro Ähre wird die Gesamtkornzahl berechnet.
Bei der Abbildung der Kornfüllung wird von einer maximalen Einspeicherungsrate pro Korn ausgegangen. Dieser Anspruch erfüllt sich nach Möglichkeit aus der täglichen Wachstumsrate. Sollte ein Defizit verbleiben, wird dieses aus den vorhandenen Assi- milatvorräten bzw. durch Umlagerung aus der grünen Biomasse beseitigt.
Da das Modell TRITSIM für limitierte Wasser- und Stickstoffversorgung entwickelt wurde, finden in den einzelnen Wachstums- und Entwicklungsprozessen (Tab. 1) entsprechende Abhängigkeiten Berücksichtigung. Grundlage für die Abschätzung des Wasserversor- gungszustandes bildet der Quotient aus aktueller zu potentieller Evapotranspiration (AET/PET). Die Größen AET und RET werden neben den täglichen schichtweisen Wasservorräten auf der Grundlage des Mehrschicht-Bodenwassermodells BOWA nach Koitzsch (Koitzsch, 1977; Koitzsch et al., 1980; Mirschel et ai, 1991) berechnet. Beim Wasser wird eine Kurzzeit- und eine sich nur zwischen DC 30 und DC 39 herausbildende Langzeit-Streßwirkung berücksichtigt. Während die erste negativ auf alle Wachstums- und Entwicklungsprozesse wirkt, beeinflußt die zweite den Prozeß der Photosynthese positiv.
Letzteres läßt sich dadurch erklären, daß Biomasse, die unter länger anhaltendem Trockenstreß gebildet worden ist, mehr aktive Spaltöffnungen pro Einheit grüner Biomas- se aufweist. Dieses Streßkompensationswachstum wurde bereits von Hsiao etal. (1976) nachgewiesen.
Der sich auf einzelne Prozesse auswirkende Stickstoffversorgungszustand des Bestandes wird über die mögliche N-Aufnahme und N-Einlagerung in die Pflanzen im Vergleich mit einem ontogeneseabhängig-notwendigen N-Gehalt in den Weizenpflanzen bestimmt. Die Grundidee des verwendeten Algorithmus geht auf Seligman et al. (1975) zurück.
Innerhalb des Modells existieren Schnittstellen, über die Modelle für die tierischen Schädlinge Getreideblattläuse und Getreidehähnchen und die Pflanzenkrankheit Wei- zenmehltau angekoppelt werden können. Die Wechselwirkungen zwischen den Scha- derregern und dem Kulturpflanzenbestand sind ausführlich bei Ebert et al. (1986) be- schrieben.
Die Wirkungen der drei abgebildeten Schaderreger werden in sogenannte Primär- und Sekundärwirkungen unterteilt. Die Primärwirkungen betreffen direkte Stoffverluste (Ver- luste durch das Fressen der Getreidehähnchen an den grünen Blättern, Verluste an für die Kornfüllung bestimmten Assimilaten durch das Absaugen von Phloemsaft durch die Getreideblattläuse und Verluste an Assimilaten durch den Assimilatverbrauch des Mehltaus). Die Sekundärwirkungen betreffen Rückkopplungen auf die Wachstums- und Entwicklungsprozesse des Weizenbestandes (Bestandesaufhellung durch die Fraßstellen der Getreidehähnchen, Einschränkung des Gasaustausches an den Stellen von Honigtau- ausscheidungen durch die Läuse, beschleunigte Seneszenz an den Mehltaukrankheits- herden).
In Tab. 1 sind die qualitativen funktionalen Abhängigkeiten der wichtigsten Teilprozesse zusammenfassend dargestellt.
Tab. 1: Übersicht über die qualitativen funktionalen Zusammenhänge der wichtigsten Teilprozesse in TRITSIM
Pflanzen bestand:
Ontogenese (ON) Photosynthese/
Assimilatbildung (A) Erhaltungs- und
Wachstumsatmung (EWA) grüne Biomasse (GBM) Kornbiomasse (KBM) Wurzelbiomasse/
Wurzeldichte (WBM) Seneszenz-Biomasse (SBM) N-Entzug Pflanze (NEP) Ährenzahl (ÄZ)
Kornzahl (KZ)
Blattflächenindex (LAI) Transpiration (TR) Bestandeshöhe (BH) Exsudatausscheidung (EX) Wurzeltiefe (WT)
Boden:
Bodentemperatur (BT) Bodenwasservorrat (BWV) N-Mineralisierung (NM) Bodenstickstoffvorrat (BNV)
= /(T, BWV, BNV, ON)
= f (ST, T, GBM, WBM, A, BWV, BNV, TL, GH, GBL, WMT, ON)
= f (A, GBM, KBM, WBM, T, TL, ON)
= /(A, ON, GH, GMT, GBM)
= /(A, ON, KZ, GBL)
= f(A, ON, WBM, BWV, BT)
= f (ON, BWV, BNV, GBM, KBM, NEP, WMT, SBM)
= f (KBM, GBM, WBM, BNV, NEP, ON)
= /(GBM, ON)
= /(ÄZ, ON, BWV, BNV, KZ)
= /(ON, GBM)
= f (BWV, T, W, RLF, ST, LAI, ON)
= f(ON)
= f(WBM, EX)
= f(ON)
= /(T, BH)
= /(T, N, ST, RLF, W, ON, BH)
= /(BWV, BT, NPOT)
= /(DÜ, NM, BNV, IM, NEP)
Dabei bedeuten: GH - Getreidehähnchen
GBL - Getreideblattlaus WMT - Weizenmehltau DU - Stickstoff-Düngung T - Temperatur
ST - Strahlung NS - Niederschlag TL - Tageslänge W - Windgschwindigkeit RLF - relative Luftfeuchte
IM - trockene und feuchte Stickstoffimmission NPOT - potentiell mineralisierbarer Bodenstickstoff
Eine ausführliche mathematische Darstellung der Algorithmen ist bei Matthäus et al.
(1986), Mirschel et al. (1987) und Matthäus et al. (1991) zu finden.
3.2 Simulationsergebnisse
Das Modell liegt in einer Version für den Ökosystem-Simulator SONCHES (Wenzel etal., 1986; Wenzel et a/., 1989) vor. Alle Simulationsrechnungen wurden unter SONCHES realisiert.
Es werden Simulationsergebnisse der Anbaujahre 1986/87 bis 1991/92 für die beiden Brandenburger Standorte Müncheberg und Hohenfinow vorgestellt.
Beide Standorte unterscheiden sich aus meteorologischer Sicht nur geringfügig (mittlere Jahrestemperatur: 8.2°C (Müncheberg), 8.2°C (Hohenfinow); mittlerer Jahresniederschlag:
533 mm (Müncheberg), 550 mm (Hohenfinow)). Während der sandige Boden am Standort Müncheberg eine Feldkapazität von 140 mm (90 cm Tiefe) aufweist, hat der Boden in Hohenfinow eine Feldkapazität von 175 mm (90 cm Tiefe). Es kamen die Winterweizen- sorten "Alcedo" und "Faktor" zum Anbau.
Für die drei Anbaujahre in Müncheberg wurden für je eine beregnete und eine unbe- regnete Variante Simulationsrechnungen durchgeführt, so daß sich insgesamt sieben Jahresreihen ergeben.
Die Ergebnisvorstellung beschränkt sich auf die dynamischen Pflanzenzustandsgrößen Ontogenese und Biomasse.
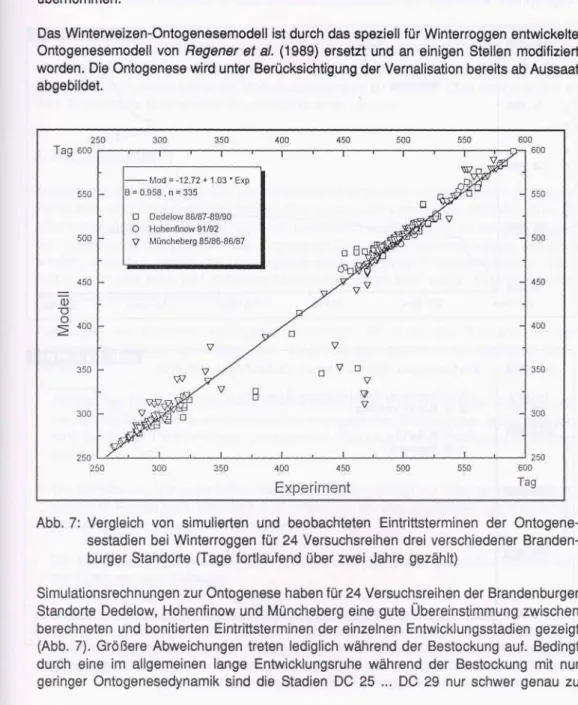
Da die Ontogenese im Winterweizenmodell als Zeitsteuerung fungiert, muß sie besonders genau modelliert werden. In Abb. 3 ist ein Modell-Experiment-Vergleich über alle Jahre, Varianten und Ontogenesestadien dargestellt. Gegenübergestellt werden die berechneten und beobachteten Eintrittstermine (dargestellt als laufender Kalendertag (KT)) der Entwicklungsstadien. Die ausgefüllten Punkte markieren dabei die wichtigen Stadien Schossen (DC 31), Blüte (DC 61) und Todreife (DC 92). Die Anpassungsgüte wurde mit Hilfe einer linearen Regressionsgleichung eingeschätzt. Ein statistischer Test zwischen der Quadrantenhalbierenden und der Regressionsgeraden ergab bei 5 % Irrtumswahr- scheinlichkeit keinen signifikanten Unterschied. Größere Abweichungen gab es in einzel- nen Jahren bei der richtigen Abschätzung der einzelnen Bestockungsstadien, denn meistens hatte der Zeitraum der Bestockung bereits im Herbst begonnen und erstreckte sich mit Ruhepausen über den Winter hinweg.
Die mittlere absolute Abweichung beträgt über alle Jahre und Stadien 6.5 Tage. Be- trachtet man nur die Stadien Schossen und Blüte, sind das im Mittel 6.0 bzw. 3.5 Tage.
Ein wichtiger Termin ist das Entwicklungsstadium Blühbeginn, da mit Erreichen von DC 61 der Prozeß der Kornfüllung aktiviert wird. Eine ungenaue Abschätzung der Länge der Kornfüllungsperiode hat große Auswirkungen auf den simulierten Kornertrag. Wie experi- mentell für Bedingungen des Standortes Müncheberg ermittelt wurde, bedeutet eine Verkürzung der Kornfüllungsperiode um einen Tag eine Reduzierung des Kornertrages um 1 - 2 dt/ha.
245-
KTModell
215-
175-
135-
95-
55
340
300-
O = 1906/87 Müncheberg a = 1987/88 Müncheberg o - 1988/89 Müncheberg A = 1991/92 Hohenfinow
300 340 135 175 215 245
^Experiment
Abb. 3: Vergleich von simulierten und beobachteten Eintrittsterminen einzelner Onto- genesestadien bei Winterweizen [KT - laufender Kalendertag] (ausgefüllte Punkte markieren DC 31 [Schossen], DC 61 [Blüte] und DC 92 [Todreife]) Für den gesamten Nährstoffkreislauf innerhalb eines Agroökosystems ist die oberirdisch gebildete Biomasse wichtig, da sie durch Weiterverwendung im landwirtschaftlichen Betrieb dem System entzogen wird. Gebunden an diese Biomasse ist natürlich auch eine bestimmte Stickstoffmenge, die damit gleichfalls das System verläßt. Eine Gegenüber- stellung der simulierten und im Experiment bestimmten oberirdischen Gesamtbiomasse ist in Abb. 4 wiedergegeben. Daraus wird ersichtlich, daß das Modell die oberirdische Biomasse bis April/Mai unter- und zur Ernte hin in der Regel geringfügig überschätzt.
Letzteres erscheint plausibel, werden doch bei der experimentellen Bestimmung der Biomassen die bereits vertrockneten und abgefallenen Pflanzenteile (meistens die untersten Blätter) nicht mehr mit erfaßt. Über alle Jahre betrachtet wird die oberirdische Biomasse modellmäßig leicht unterschätzt, wie die Koeffizienten der entsprechenden Regressionsgleichung zeigen. Die mittlere relative Abweichung zwischen Modell und
Experiment beträgt bei der oberirdischen Biomasse über alle Meßzeitpunkte (Zwischen- ernten) ca. 30 % .
[dt/hcü 180
160-
140-
120-
GBMMo r-5,61+1,02 GBMEx
B = 0,889 n = 64
O = 1986/87 Müncheberg D = 1987/88 Müncheberg o = 1988/89 Müncheberg A - 1991/92 Hohen finow
A CD
100 120 no 160 180 200 GBMExperiment Cdt/ha]
Abb. 4: Vergleich der für Winterweizen simulierten (GBMModell) und gemessenen (GBMExperiment) gesamten oberirdischen Biomasse
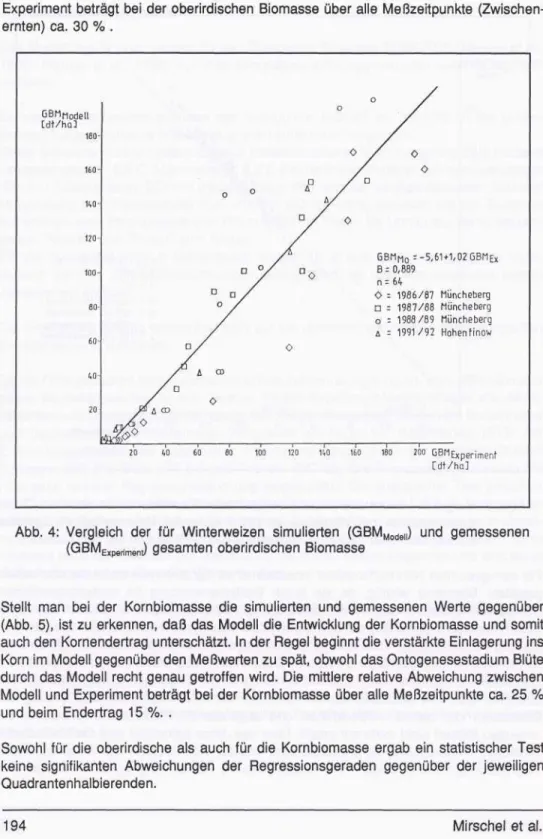
Stellt man bei der Kornbiomasse die simulierten und gemessenen Werte gegenüber (Abb. 5), ist zu erkennen, daß das Modell die Entwicklung der Kornbiomasse und somit auch den Kornendertrag unterschätzt. In der Regel beginnt die verstärkte Einlagerung ins Korn im Modell gegenüber den Meßwerten zu spät, obwohl das Ontogenesestadium Blüte durch das Modell recht genau getroffen wird. Die mittlere relative Abweichung zwischen Modell und Experiment beträgt bei der Kornbiomasse über alle Meßzeitpunkte ca. 25 % und beim Endertrag 15 %. .
Sowohl für die oberirdische als auch für die Kornbiomasse ergab ein statistischer Test keine signifikanten Abweichungen der Regressionsgeraden gegenüber der jeweiligen Quadrantenhalbierenden.
KBMMo:: 1,10 +0,863 B = 0,802
n = 25
O = 1986/87 Müncheberg a = 1987/88 Müncheberg o = 1988/89 Müncheberg A = 1991/92 Hohenfinow
20 40 60 80 100 120 KBMExperiment Cdt/ha]
Abb. 5: Vergleich der für Winterweizen simulierten (KBMModell ) und gemessenen (KBMExperiment) Kornbiomasse [ausgefüllte Punkte markieren den Endernteer- trag]
In Abb. 6 werden als Beispiel für die Jahre 1987/1988 (Müncheberg, beregnet) und 1991/1992 (Hohenfinow) die mitTRITSIM simulierten oberirdischen Gesamt- und Kornbio- massen im Vergleich mit den Meßwerten dargestellt.
2800.000
1500.000-
1000.000 B
i o n a s s
e 500.000-
Uinterueizen (Sorte Faktor) CMUncheberg 1987X1988, beregnet]
1 — gesamte oberirdische Biomasse
2 — Kornbiomasse •*•
Modell + fiep wer t
8-Oct 10-Dec 12-Feb 16-Apr 19-Jui 22-Aug
1600.000 [g/m23 1200.800 B
i
0 880.000 m
a s
e 400.000
0.000
nade by SONCHES Uinterneizen (Sorte Alcedo) LHohenfinow 1991/19923
1 — gesamte oberirdische Biomasse 2 — Kornbiomasse
Modell
* Meßwert
20-Oct 20-Dec 19-Feb 21-Apr 21-Jun 22-Aug Abb. 6: Modell-Experiment-Vergleich für die Dynamik der gesamten oberirdischen sowie
der Kornbiomasse für die Anbaujahrel987/1988 (Müncheberg, beregnet) und 1991/1992 ( Hohenfinow) für Winterweizen
4. Wachstums- und Ertragsbildungsmodell für Winterroggen
Aufgrund der Verwandtschaft der Wintergetreidearten liegt es nahe, die Modellstrukturen und -algorithmen für den Winterweizen auch für die Modellierung anderer Getreidearten zu nutzen. Daß dies erfolgsversprechend ist, zeigen die von Wechsung et al. (1992) auf der Grundlage der TRITSIM-Strukturen und TRITSIM-Modellalgorithmen durchgeführten Modellierungsarbeiten für Sommergerste.
Erste Ergebnisse der Übertragung der Modellstrukturen von TRITSIM auf Winterroggen sollen im folgenden vorgestellt werden.
Das generelle Modellkonzept von TRITSIM (Triebkräfte, Kompartimentierung, Kohlenstoff-, Wasser- und Stickstoffbilanzierung, Kopplung Boden - Pflanze) wurde unverändert übernommen.
Das Winterweizen-Ontogenesemodell ist durch das speziell für Winterroggen entwickelte Ontogenesemodell von Regener et al. (1989) ersetzt und an einigen Stellen modifiziert worden. Die Ontogenese wird unter Berücksichtigung der Vernalisation bereits ab Aussaat abgebildet.
Tag eoo
450 500 550
— Mod = -12.72 + 1.03*Exp B = 0.958, n = 335
D Dedelow 86/87-89/90 O Hohenfinow 91/92 V Müncheberg 85/86-86/87
450
T50 O
300
250
300 350
Experiment
Abb. 7: Vergleich von simulierten und beobachteten Eintrittsterminen der Ontogene- sestadien bei Winterroggen für 24 Versuchsreihen drei verschiedener Branden- burger Standorte (Tage fortlaufend über zwei Jahre gezählt)
Simulationsrechnungen zur Ontogenese haben für 24 Versuchsreihen der Brandenburger Standorte Dedelow, Hohenfinow und Müncheberg eine gute Übereinstimmung zwischen berechneten und bonitierten Eintrittsterminen der einzelnen Entwicklungsstadien gezeigt (Abb. 7). Größere Abweichungen treten lediglich während der Bestockung auf. Bedingt durch eine im allgemeinen lange Entwicklungsruhe während der Bestockung mit nur
berechnen. Im Modell werden diese Stadien in der Regel zu früh erreicht. Dieser Fakt ist verantwortlich für die in Abb. 7 deutlich unterhalb der Quadrantenhalbierenden liegenden Punkte. Andererseits ist dies aber ohne Auswirkung auf die Genauigkeit der Berechnung des Schoßbeginns (DC 30) sowie darauf folgender Ontogensestadien. Die Eintrittstermine der Entwicklungsstadien ab Schossen werden durch das Modell in der Regel um einige Tage zu spät erreicht.
188.888-1
0 75.888- n
t o
9 58.888
e n e s
e 25.888
Uinterroggen (Sorte Pluto) CHohenfinow 1991/92]
Modell + Meßwert
8.888
22-Sep 28-No«J 3-Feb 18-Apr 16-Jum 22-Aug
Uinterroggen (Sorte Pluto) CHohenfiuou 1991/923 1 = gesamte oberirdische Biomasse
2 = Kornbicnasse Modell Meßwert
Abb. 8: Simulierte Ontogense und Biomassekomponenten im Vergleich zu gemessenen Werten für Winterroggen [Standort Hohenfinow, 1991/1992]
Da die Ontogenese im Ertragsbildungsmodell als Zeitsteuergröße fungiert, ist es not- wendig, daß die Eintrittstermine bestimmter Stadien möglichst genau getroffen werden, wie z.B. Schossen, Blüte und Reife. Die durchschnittlichen absoluten Abweichungen über alle Versuchsreihen betragen für das Schossen (DC 30) 3.9 Tage, für die Blüte (DC 61) 4.5 Tage und für die Reife (DC 91/92) 5.8 Tage. Abb. 8 enthält für die Ontogenese einen Modell-Experiment-Vergleich für den Standort Hohenfinow 1991/92.
Die Parameter der Wachstums- und Ertragsbildungsprozesse wurden aufgrund experi- menteller Daten der oben genannten Standorte rekalibriert. Simulationsstudien haben allerdings gezeigt, daß eine bloße Parameterneuschätzung nicht für alle Prozesse ausreicht, um zufriedenstellende Modellanpassungen zu erzielen. Dies wird aus den in Abb. 8 gleichfalls dargestellten Biomassefraktionen deutlich.
5. Modellreduktion
Anwendung und Übertragbarkeit von Agroökosystemmodellen erfordern einen erfüllbaren Parameter- und Eingangsgrößenbedarf. Das ist bei sehr detaillierten Modellen aber oft nicht realisierbar. Deshalb sind Fragen der praktischen Modellnutzung immer auch Fragen der Modellreduzierung. Kann die Eingangsgrößen- und Parametermenge reduziert werden, ohne daß wesentliche Genauigkeitsverluste eintreten ? Inwieweit das für TRIT- SIM möglich sein kann, soll in diesem Abschnitt für einige sehr starke Vereinfachungen von Prozessen untersucht werden.
Abgesehen von kleineren Änderungen technischer Art wurde die Grundstruktur des Kulturpflanzenmodells beibehalten. Die Veränderungen betreffen die folgenden Kom- ponenten:
• Anstelle des Mehrschicht-Bodenwassermodells tritt ein Einschicht-Bodenwassermodell mit dynamisch dem Wurzeltiefenwachstum angepaßter Schichtdicke. Jeder Standort wird durch das Parameterpaar permanenter Welkepunkt und Feldkapazität charak- terisiert.
• Die Berechnung der potentiellen Evapotranspiration erfolgt mit Hilfe einer ganzjährig nutzbaren Formel nach Wendung etal. (1991), die als Eingangsgrößen nur die Global- strahlung und die Tagesmitteltemperatur erfordert.
• Die Bodentemperatur wird Standort- und tiefenabhängig aus gleitenden Mittelwerten der Lufttemperatur geschätzt.
• Analog zur Einschichtbetrachtung des Wasserhaushaltes erfolgt die des Bodenstick- stoffes auch nur über eine Schicht. Berücksichtigt werden die Prozesse Minera- lisierung, Stickstoffentzug durch die Pflanze, atmosphärischer Stickstoffeintrag und Düngung.
Durch diese Veränderungen wurde die Abbildung des Wasser- und Stickstoffhaushaltes sehr stark vereinfacht. Der notwendige Eingangsgrößenbedarf konnte um 40 %, der Parameterbedarf konnte um ca. 20 % gesenkt werden.
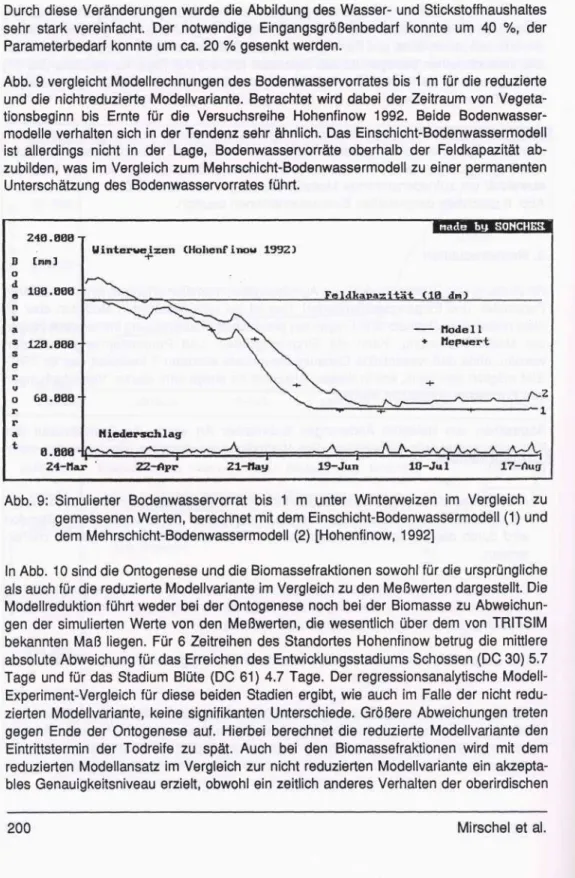
Abb. 9 vergleicht Modellrechnungen des Bodenwasservorrates bis 1 m für die reduzierte und die nichtreduzierte Modellvariante. Betrachtet wird dabei der Zeitraum von Vegeta- tionsbeginn bis Ernte für die Versuchsreihe Hohenfinow 1992. Beide Bodenwasser- modelle verhalten sich in der Tendenz sehr ähnlich. Das Einschicht-Bodenwassermodell ist allerdings nicht in der Lage, Bodenwasservorräte oberhalb der Feldkapazität ab- zubilden, was im Vergleich zum Mehrschicht-Bodenwassermodell zu einer permanenten Unterschätzung des Bodenwasservorrates führt.
240.808 n [mml 188.808-
s 128.888-
68.808-
U interne izen (Hohenfinow 1992)
Feldkapazität (10 dm) Modell Hepwert
Niederschlag
_>\ s^ A^yv A
24-Mar 21-llay 19-Jun 10-Jul 17-rtug
Abb. 9: Simulierter Bodenwasservorrat bis 1 m unter Winterweizen im Vergleich zu gemessenen Werten, berechnet mit dem Einschicht-Bodenwassermodell (1) und dem Mehrschicht-Bodenwassermodell (2) [Hohenfinow, 1992]
In Abb. 10 sind die Ontogenese und die Biomassefraktionen sowohl für die ursprüngliche als auch für die reduzierte Modellvariante im Vergleich zu den Meßwerten dargestellt. Die Modellreduktion führt weder bei der Ontogenese noch bei der Biomasse zu Abweichun- gen der simulierten Werte von den Meßwerten, die wesentlich über dem von TRITSIM bekannten Maß liegen. Für 6 Zeitreihen des Standortes Hohenfinow betrug die mittlere absolute Abweichung für das Erreichen des Entwicklungsstadiums Schossen (DC 30) 5.7 Tage und für das Stadium Blüte (DC 61) 4.7 Tage. Der regressionsanalytische Modell- Experiment-Vergleich für diese beiden Stadien ergibt, wie auch im Falle der nicht redu- zierten Modellvariante, keine signifikanten Unterschiede. Größere Abweichungen treten gegen Ende der Ontogenese auf. Hierbei berechnet die reduzierte Modellvariante den Eintrittstermin der Todreife zu spät. Auch bei den Biomassefraktionen wird mit dem reduzierten Modellansatz im Vergleich zur nicht reduzierten Modellvariante ein akzepta- bles Genauigkeitsniveau erzielt, obwohl ein zeitlich anderes Verhalten der oberirdischen
Gesamtbiomasse zu erkennen ist. Bedingt durch die im reduzierten Modellansatz ver- längerte Kornfüllungsperiode wird der Endertrag überschätzt.
Dieser erste Versuch zeigt, daß es für bestimmte Fragestellungen möglich sein kann, Modellstrukturen zu vereinfachen und damit den Umfang von Eingangsgrößen und Para- metern merklich zu verringern, ohne wesentliche Genauigkeitsverluste hinnehmen zu müssen.
188.888 CDCD
0 75.888- n
t o
9 58.886-
e 25.888-
8.888 H
Winterueizen (Hohenfinow 1992)
... ohne M o d e l I r e d u k t i o n nit Modellreduktion + Meßwert
24-Mar 22-flpr 21-May 19-Jun 18-Jul 17-flug
2000.000 n
1588.000- B
i o
M1008.000-
508.888-
Ninterweizen (Hohenfinou 1992)
... ohne M o d e l I r e d u k t i o n M i t Mode lIreduktion + Meßwert
oberirdisch
8 . 8 8 8 1
24-Mar 22-flpr 21-May 19-Jun
1 1 18-Jul 17-flug
Abb. 10: Mit reduziertem und nicht reduziertem Modellansatz simulierte Ontogense und Biomassekomponenten im Vergleich zu gemessenen Werten [Hohenfinow, 1992]
6. Literatur
Bellmann, K.; Ebert, W.; Freier, B.; Kunkel, K.; Matthäus, E.; Schuitz, A.; Wenzel, V., 1986. Agroecosystem modelling and Simulation - The winter wheat agroecosystem model AGROSIM-W. In: Tag.-Ber. 242, Akad.Landwirtsch.Wiss. DDR. Berlin, S. 5-28 Ebert, W.; Matthäus, E.; Schuitz, A., 1986. Use of the agroecosystem model winter wheat (AGROSIM-W) for assessment of complex infestation situations. In: Tag.-Ber.
242, Akad.Landwirtsch.Wiss. DDR. Berlin, S. 113-127
Hsiao, T.C.; Fereres, E.; Acevedo, E.; Henderson, D.W., 1976. Water stress and dynamics of growth and yield of crop plants. In: LANGE, O.L; KAPPEN, L; SCHULZE, E.D.: Water and plant life, Ecological Studies 19, Berlin-West, S.281-305
Kartschall, T.; Matthäus, E.; Asseng, S., 1990. Simulation Experiments with the Agroe- cosystem Model DEMETER Using SONCHES. Syst. Anal. Model. Simul. 7, S. 429 - 438
Koitzsch, R., 1977. Schätzung der Bodenfeuchte aus meteorologischen Daten, Boden- und Pflanzenparametern mit einem Mehrschichtenmodell. Z. Meteorologie 27, S. 302- 306
Koitzsch, R.; Helling, R.; Vetterlein, E., 1980. Simulation des Bodenfeuchteverlaufs unter Berücksichtigung der Wasserbewegung und des Wasserentzuges durch Pflan- zenbestände. Arch.Acker- Pflanzenbau Bodenkd.24, S.717-725
Leser, H., 1991. Landschaftsökologie. Eugen-Ulmer-Verlag, Stuttgart, Uni-Taschenbuch Bd. 521
Lutze, G.; Schuitz, A.; Wenkel, K.-O., 1993. Vom Populationsmodell zum Landschafts- modell - Neue Herausforderungen und Wege zur Nutzung von Modellen in der Agrar- landschaftsforschung. Z. f. Agrarinformatik 1, S. 19-25
Matthäus, E.; Kartschall, T.; Frühauf, C.; Grossmann, S.; Wechsung, F., 1991. The Agroecosystem model winter wheat DEMETER. Potsdam-Institut für Klimafolgenfor- schung, Eigenverlag, 61 S.
Matthäus, E.; Knijnenburg, A.; Schäfer, W., 1984. A winter wheat crop production model on the basis of carbon dioxide measurements under field conditions. Wiss. Z.
Humboldt Univ. (Mathem.-naturwiss. Reihe) 33, S. 362-368
Matthäus, E.; Mirschel, W.; Kretschmer, H.; Kunkel, K.; Klank, L, 1986. The winter wheat crop model TRITSIM of the agroecosystem AGROSIM-W. In: Tag.-Ber. 242, Akad.Landwirtsch.Wiss. DDR. Berlin, S. 43-74
Mirschel, W.; Kretschmer, H.; Klank, I.; Matthäus, E.; Kunkel, K., 1987. Dynamisches Ertragsbildungs- und Entwicklungsmodell TRITSIM für Wint9rweiz9n 1. Mitteilung:
Modellbeschreibung. Arch. Acker- Pflanz9nbau Bodenkd. 31, S 249-259
Mirschel, W.; Kretschmer, H.; Matthäus, E.; Koltzsch, R., 1991. Simulation of the effect of nitrogen supply on yield formation processes in winter wheat with the model TRIT- SIM. Fertilizer Research 27, S. 293-304
Regener, E.; Reuter, G., 1989. Entwicklung eines dynamischen Ertragsbildungsmodells für Winterroggen. Forschungs-Bericht, IPZ Gülzow
Seligman, N.G.; Keulen, H. van; Goudriaan, J., 1975. An elementary model of nitrogen uptake and redistribution by annual plant species. Oecologia 21, S. 243-261
Wechsung, F.; Waloszczyk, K.; Bauer, M.; Gräfe, J., 1992. Possibilities of Use of the Model System DEMETER-TRITSIM for Construction of Dynamic Cereal Models Exemplary of Spring Barley. In: Proceedings of the International Congress on Agro- Ecosystem Modelling (5th-9th October 1992, Braunschweig), S. 108
Weir, A.U.; Bragg, P.L.; Porter, J.R.; Rayner, J.H., 1984. A winter wheat crop Simulation model without water or nutrient limitations. J. agric. Sei. 102, S.371-382
Wendung, U.; Schellin, H.-G.; Thomä, M., 1991. Bereitstellung von täglichen Infor- mationen zum Wasserhaushalt des Bodens für die Zwecke der agrarmeteorologischen Beratung. Z. f. Meteorologie 41
Wenzel, V.; Bellmann, K.; Matthäus, E., 1989. Rechnergestützte Modellierung mittels SONCHES. In: Tag.-Ber. 275, Akad.Landwirtsch.Wiss. DDR. Berlin, S. 313-319 Wenzel, V.; Matthäus, E.; Flechsig, M., 1986. The Simulation System SONCHES. In:
Tag.-Ber. 242, Akad.Landwirtsch.Wiss. DDR. Berlin, S. 29-42

![Abb. 3: Vergleich von simulierten und beobachteten Eintrittsterminen einzelner Onto- Onto-genesestadien bei Winterweizen [KT - laufender Kalendertag] (ausgefüllte Punkte markieren DC 31 [Schossen], DC 61 [Blüte] und DC 92 [Todreife]) Für den gesamten Nährs](https://thumb-eu.123doks.com/thumbv2/1library_info/4604406.1604033/11.727.53.627.89.587/vergleich-simulierten-beobachteten-eintrittsterminen-genesestadien-winterweizen-kalendertag-ausgefüllte.webp)
![Abb. 5: Vergleich der für Winterweizen simulierten (KBM Modell ) und gemessenen (KBM Experiment ) Kornbiomasse [ausgefüllte Punkte markieren den Endernteer-trag]](https://thumb-eu.123doks.com/thumbv2/1library_info/4604406.1604033/13.718.52.647.84.754/vergleich-winterweizen-simulierten-gemessenen-experiment-kornbiomasse-ausgefüllte-endernteer.webp)
![Abb. 8: Simulierte Ontogense und Biomassekomponenten im Vergleich zu gemessenen Werten für Winterroggen [Standort Hohenfinow, 1991/1992]](https://thumb-eu.123doks.com/thumbv2/1library_info/4604406.1604033/16.724.90.679.74.913/simulierte-ontogense-biomassekomponenten-vergleich-gemessenen-winterroggen-standort-hohenfinow.webp)
![Abb. 10: Mit reduziertem und nicht reduziertem Modellansatz simulierte Ontogense und Biomassekomponenten im Vergleich zu gemessenen Werten [Hohenfinow, 1992]](https://thumb-eu.123doks.com/thumbv2/1library_info/4604406.1604033/19.721.71.641.84.826/reduziertem-reduziertem-modellansatz-simulierte-ontogense-biomassekomponenten-vergleich-hohenfinow.webp)
