www.nul‐online.de 1
Anhänge zur Veröffentlichung:
FRETZER, S. (2015): Vorstellung einer ökosystemaren FFH‐
Verträglichkeitsprüfung – ein Modell anhand relevanter Schlüsselarten für das Nahrungsnetz.
Naturschutz und Landschaftsplanung 47 (10), 319‐327.
In diesem Anhang sind alle Ecopath-, Ecosim- und Ecospace-Einstellungen enthalten, die für die Replikation der Ergebnisse unter Verwendung der Ecopath-Software (www.ecopath.org) notwendig sind. Die Ergebnisse beruhen auf dem Ecopath-Modell, welches zur Untersuchung der Fachkonventionen entwickelt wurde und das Ökosystem vor dem Bau eines Industriegebiets analysiert (siehe FRETZER&MÖCKEL 2014).
Am Schluss befinden sich noch alle farbigen Abbildungen des Manuskripts und eine Liste mit allen Literaturquellen.
Inhalt des Anhangs Inhalt
1. Ecopath ... 1
2. Ecosim ... 7
3. Ecospace ... 11
4. Sensitivitätsanalyse ... 12
5. Abbildungen in Farbe ... 14
Referenzen: ... 18
1. Ecopath
Ecopath ist eine quantitative Modellierungsmethode, die die Biomasse-Flüsse zwischen funktionellen Gruppen beschreibt (CHRISTENSEN et al. 2008). Eine funktionelle Gruppe kann aus einer einzelnen Art oder Population bestehen oder sich auf einzelne oder mehrere taxonomische Familien oder Taxa beziehen, zum Beispiel kann eine funktionelle Gruppe
‚Hirschkäfer (Lucanus cervus)‘ nur eine Art beschreiben oder die funktionelle Gruppe könnte auch ‚Käfer (Coleoptera)‘ zusammengefasst darstellen. Die Größe einer funktionellen Gruppe oder z.B. einer Population kann entweder durch die Individuenzahl oder durch die Biomasse gemessen werden (WEHNER & GEHRING 1995). Für jede funktionelle Gruppe wird angenommen, dass innerhalb eines definierten Zeitraums Input und Output, also Produktion und Verbrauch einer Gruppe gleich sind (CHRISTENSEN et al. 2008).
Um die grundlegenden Matrixberechnungen in Ecopath durchführen zu können, wird für jede funktionelle Gruppe i die Gleichung wie folgt dargestellt (CHRISTENSEN et al. 2008):
www.nul‐online.de 2 0
) / ( )
/ (
1
i i i
n
j
ji j j
i i
i P B EE B Q B DC Y E BA
B
wobei Bi die Biomasse der Beutegruppe i und Bj die Biomasse des Räubers j darstellt, (P/B)i ist die Produktion/Biomasse-Rate für die funktionelle Gruppe i, EEi ist die ökotrophische Effizienz, die den Anteil der Produktion beschreibt, die tatsächlich im Ökosystem genutzt wird, (Q/B)j ist die Konsumption/ Biomasse-Rate des Räubers j, DCij beschreibt den Anteil, den die Beutegruppe i im Nahrungsspektrum von Räuber j darstellt, Yi ist die gesamte Fang-oder Jagdquote bezüglich der funktionellen Gruppe i, Ei ist die Netto-Migrationsrate der Gruppe i, BAi ist die Biomasse-Akkumulationsrate der Gruppe i und n ist die Anzahl der funktionellen Gruppen im Model.
Die Daten, die in Ecopath verwendet werden bestehen aus einer Reihe Basisparameter, die für jede funktionelle Gruppe für einen definierten Zeitraum bestimmt werden müssen: Biomasse (B in t km-²), Produktion/Biomasse-Rate (P/B, pro Jahr), welches der Gesamtsterblichkeitsrate (Z) entspricht; Konsumption/Biomasse-Rate (Q/B, pro Jahr); und die ökotrophische Effizienz EE (CHRISTENSEN et al. 2008). Falls einer dieser Parameter für eine funktionelle Gruppe nicht ermittelt werden kann, ist es notwendig Ecopath zusätzliche Daten zur Verfügung zu stellen, sodass die Software die fehlenden Parameter berechnen kann, z.B. Nettomigrationsrate, Fangquoten, Biomasseakkumulationsrate und Assimilationsrate (CHRISTENSEN et al. 2008).
Die Produktion einer funktionellen Gruppe bezieht sich auf die Produktion von Gewebe bzw.
neuer Biomasse, unabhängig davon ob diese überlebt oder in den Detrituspool fließt.
(CHRISTENSEN et al. 2008).
Folgende Parameter und Indices wurden für die Untersuchung des Models herangezogen (nach (CHRISTENSEN et al. 2008):
Trophieebene TE: gibt die Ernährungsstufe der funktionellen Gruppe wieder, Höhe der Stufe ist abhängig von den Trophiebenen der Beutegruppen
P/B-Rate: reflektiert die Gesamtsterblichkeitsrate einer funktionellen Gruppe zusammensetzend aus natürlicher, durch Räuber bedingter- und durch Jagd bedingter Sterblichkeitsrate;
Gesamtdurchsatz des Systems
(Total System Throughput TST):
beschreibt die Summe aller Energieflüsse im System (in t km-2Jahr-1):
Gesamtdurchsatz (TST) = ∑Konsumption + ∑Export +∑Respiration + ∑Fluss in den Detritus-Pool
Die Größe eines Ökosystems wird nicht durch die Biomasse, sondern den Gesamtdurchsatz verdeutlicht.
Verbindungs-Index (Connectance Index):
berechnet die Anzahl der tatsächlichen
Nahrungsverbindungen im System in Relation zu der Anzahl aller möglichen Verbindungen
Omnivorie Index (OI): wird berechnet nach der Varianz der Trophieebenen der Beutegruppen, die vom Räuber gefressen werden; ein hoher Index zeigt an, dass der Räuber Beutegruppen über
verschiedene Trophieebenen konsumiert, während 0 einen Nahrungsspezialisten kennzeichnet
www.nul‐online.de 3 Modelstruktur und Dateninput
Die funktionellen Gruppen und Parameter des Waldgebietes basieren auf den zoologischen (DOROW et al. 2004) und waldkundlichen (KEITEL & HOCKE 1997) Untersuchungen des Naturreservats Schönbuche in Hessen, bei dem es sich um zwei Wald-Teilgebiete mit einem Hainsimsen-Buchenwald handelt. Die Daten für die Ackerfläche wurden aus der Literatur übernommen, wobei zwei verschiedene Ackerfrüchte gewählt wurden (Tab. 1A im Anhang).
Grasflächen gelten als bevorzugtes Nahrungsfläche des Rotmilan (MOUGEOT et al. 2011;
KNOTT et al. 2009), da aus Kapitel F keine genauen Angaben zur Landwirtschaft vor Ort zu finden waren und die Abbildung eine Ackerfläche zeigt (LAMBRECHT & TRAUTNER 2007), wurden hier zwei verschiedene Szenarien berücksichtigt, Gras- und Getreideanbau. Alle Literaturangaben, die für das Ecopath-Model verwendet wurden, sind in Tabelle 1A im Anhang gelistet.
Es wurden verschiedene Modelle herangezogen, um die Parameter einzelner funktioneller Gruppen bestimmen zu können. Um die P/B-Rate der kleinen Säugetiere zu errechnen wurde ein Model verwendet, welches sich auf die Körpermasse der Säuger bezieht (WESTERN 1983;
BANSE &MOSHER 1980). Für die Insektengruppen der zoologischen Untersuchungen wurde einen Längen-Gewichts-Model verwendet, zum Beispiel für Schmetterlinge (SAGE 1982). Für die Anwendung des Längen-Gewicht-Models für Insekten (SAGE 1982) wurde auch auf Online- Datenbanken zurückgegriffen, falls für einzelne Arten keine Längenangaben in der Literatur gefunden wurden (abgerufen August/ September 2013): www.britishbugs.org.uk, www.natur- in-nrw.de, www.naturespot.org.uk, www.coleo-net.de, http://bugguide.net, www.thewcg.org.uk, www.kaefer-der-welt.de, www.insektenbox.de, www.wildebijen.nl, //ameisenwiki.de, http://de.wikipedia.org; http://en.wikipedia.org, http://ukmoths.org.uk, http://www.natur-schmetterlinge.ch; http://zipcodezoo.com; www.lepiforum.de.
Für die funktionellen Pflanzengruppen wurde ein Model, um die Biomasse abschätzen zu können (SCHULZE et al. 2009).
Einige Gruppen im Model werden bejagt, was v.a. Schwarzwild, Rot- und Rehwild und Fuchs im Naturreservats Schönbuche betrifft. Die durchschnittlichen Jagdquoten (Hessisches Ministerium für Umwelt, Klimaschutz, Landwirtschaft und Verbraucherschutz, Quelle der Daten: http://verwaltung.hessen.de/irj/HMULV_Internet, Artikel: Daten und Fakten) wurden der Standardeinheit (t km-²) des Ökosystemmodels angepasst und als Wirkfaktor mit berücksichtigt.
Da es sich hier um ein fiktives Fallbeispiel handelt, ist eine Benotung der Daten mit einem Pedigree-Index (CHRISTENSEN et al. 2008) nicht möglich, aber die Datenqualität eines Ökosystemmodels des Naturreservats Schönbuche ist als hoch (0.7 von 1.0) einzustufen.
www.nul‐online.de 4
Tabelle A1: Quellenangaben der wissenschaftlichen Studien, die zur Berechnung der Parameter der funktionellen Gruppen verwendet wurden Biomasse P/B Q/BNahrungsspektrum Schwarzwild (Sus scrofa) D.NÖLLENHEIDT, PERS.MITT., GROOT BRUINDERINK & HAZEBROEK 1994,MELIS et al. 2006
BIEBER &RUF 2005, NORES et al.2008GUSTAFSSON et al.1999 GROOT BRUINDERINK &HAZEBROEK, 1994 Rotwild (Cervus elaphus) D.NÖLLENHEIDT, PERS.MITT., MYSTERUD et al.,2001LOISON &LANGVATN 1998,MILNER- GULLAND et al.,2000
HUDSON &NIETFELD 1985MATRAI et al.2004 Rehwild (Capreolus capreolus) D.NÖLLENHEIDT, PERS.MITT., ELLENBERG 1975;MYSTERUD et al.2001
JEAN-MICHEL et al. 1993ELLENBERG1975 BARANCEKOVA 2004,STORMS et al.2008 Fuchs CAVALLINI 1995,VOS 1995 HARRIS &SMITH, 1987, HEYDON & REYNOLDS2000
WEBBON et al.2004HOELGAARD 2008, JEDRZEJEWSKI &JEDRZEJEWSKI 1992 Rotmilan KNOTT et al.2009;MOUGEOT et al.2011 BLANCO et al.1990 KNOTT et al.2009;MOUGEOT et al.2011 Vögel ALATALO 1978,CARRASCAL et al.1998,DOROW et al.2004, HURLY 1992,LINDSTRÖM & KVIST 1995,MACLEOD et al. 2008,MAGRATH 1991, MORENO et al.2005,SAETHER 1987,POLO &CARRASCAL 1999
BADYAEV 1997, BOTKIN &MILLER, 1974,DESHOLM 2009, DOBSON 1987,EKMAN &ASKENMO 1986, HIRONS 1985, HOLYOAK 1971, JOHNSTON et al.1997, NILSSON &SMITH 1985,PASINELLI 2006, RICKLEFS 2000, SIRIWARDENA et al. 1998,WIEBE2006 ALATALO 1978,BECH& PRAESTENG 2004, LINDSTRÖM &KVIST 1995,WEINER & GLOWACINSKI 1975
ALDER &MARSDEN 2010,BIBBY & GREEN 1980,CLAYTON et al.1996;PONS &PAUSAS 2007,GAJDOS &KRISTIN 1997, GAJDOS &KRISTIN 1997;GREENWOOD & HARVEY 1978;HERNANDEZ 2009,JONES et al.2006,KENWARD &SIBLY 1977, MARQUISS &RAE 1994,2002;SUMMERS et al.2010,MARCHETTI et al.1998, MOREBY &STOATE 2001,NEWTON 1964, 1967,NOUR et al.1997,CLAYTON et al. 1996;PONS &PAUSAS 2007,BIBBY & GREEN 1980;GAJDOS &KRISTIN 1997; SUHONEN et al.1994,SUMMERS ET AL. 2010,SOTNAR &OBUCH 2009;TAPIA et al.2007
www.nul‐online.de 5
Kleine Säugetiere BORATYNSKI&KOTEJA 2010; GORECKI &SZAFRANEK 1981; PETRUSEWICZ et al.1972; SHORE et al.1992;SUCHOMEL &HEROLDOVA 2008,CHITTY 1952;DAAN &SLOPSEMA, 1978,CHURCHFIELD 2002; DICKMANN 1988;MCDEVITT & ANDREWS 1997;OCHOCINSKA &TAYLOR 2003,DOROW et al. 2004, ERGON et al.2004, EVANS 1973, FERNS 1979,, KROL et al.2005,LURZ & LLOYD 2000;WAUTERSet al. 2007; WAUTERS &DHONDT 1989),TOAL et al.2001; WATTS1969,LURZ &LLOYD 2000 (BANSE AND MOSHER, 1980;WESTERN,1983)(CHURCHFIELD,2002), (DAAN AND SLOPSEMA, 1978),(KROL ET AL., 2005),(PEACOCK AND SPEAKMANN,2001), (TOAL ET AL.,2001), (VIGNAULT ET AL., 1996),(ZONG ET AL., 2010)
(BERTOLINO ET AL.,2004;L.A.WAUTERS ET AL.,2007;ZONG ET AL.,2010)(BROWN, 1987;HANSSON,1971;LARSSON AND HANSSON,1977),(BROWN,1987; HANSSON,1971;TOAL ET AL.,2001), (BERTOLINO ET AL.,2004;L.A.WAUTERS ET AL.,2007;ZONG ET AL.,2010) Schmetterlinge DOROW et al. 2004,SAGE 1982SEVERNS et al. 2006 MUTHUKRISHNAN & PANDIAN1984 DOROW et al. 2004 Stechimmen DOROW et al. 2004,SAGE 1982DOROW et al. 2004 HARDER 1986,JOSENS et al.1998,PAUL &ROCES 2003
DOROW et al. 2004 HirschkäferDOROW et al. 2004,HARVEY et al. 2011; RINK 2006FREMLIN 2009; RINK 2006RINK 2006KRENN et al. 2002; RINK &SINSCH 2011; RINK 2006) Käfer DOROW et al. 2004,SAGE 1982GRIMM 1973 GRIMM 1973, SCHAUERMANN 1977DOROW et al. 2004 Wanzen DOROWet al. 2004,GLEN 1997, SAGE 1982COHEN 1985,DOROW et al. 2004REINHARDT et al. 2007 DOROW et al. 2004,GLEN 1997 Spinnentiere CHEN &WISE 1999,DOROW et al. 2004,Phillipson 1960, SCHMITZ 2005
PALANICHAMY 1985KIM et al. 2005, PALANICHAMY 1985, PHILLIPSON 1960 BILDE &TOFT 2000,DENNO et al. 2003, HALAJ &CADY 2000,PEKAR 2000
www.nul‐online.de 6
WürmerDOROW et al. 2004,THOMPSON et al. 1993LAKHANI &SATCHELL 1970CURRY &SCHMIDT 2007 Bodenvegetation KEITEL&HOCKE 1997, SCHULZE et al. 2009 CROW 1978,LONG & TURNER 1975, SCHULZE et al.2009
Bäume BARNA 2001,JOCHHEIM et al., 2009; KEITEL&HOCKE 1997, JOHNSON &RISSER 1974; WWW.HOLZHANDEL.DE (ABGERUFEN AM 30.09.2013)
GOWER et al. 2001, MUND et al. 2010,NI et al. 2001,REINERS 1972,XIAO et al. 2003, YUSTE et al. 2005
Totholz KEITEL&HOCKE 1997;WWW.HOLZHANDEL.DE(ABGERUFEN AM 30.09.2013) DetritusJONARD et al. 2006,KEITEL &HOCKE 1997,MICHALZIK et al. 2001 Acker: Gras - FloraHECTOR et al. 1999; SPEHN et al. 2005HECTOR et al. 1999 Acker: Gras - Fauna BENTON et al. 2002; CLAPHAM 2011; DENNIS et al. 2008Siehe Quellen der obigen Gruppen: kleine Säugetiere, Vögel und Insekten Acker: Getreide - Flora ANONYMOUS 2013ANONYMOUS 2013 Acker: Getreide - Fauna HOF &BRIGHT 2010; TATTERSALL et al. 2002; WINQVIST et al. 2011
Siehe Quellen der obigen Gruppen: kleine Säugetiere, Vögel und Insekten
www.nul‐online.de 7 2. Ecosim
Mathematischer Hintergrund
Ecosim basiert auf der Kopplung folgender Differenzialgleichung für jede Beutegruppe i und Räubergruppe j des Ecopath-Modells (CHRISTENSEN et al. 2008):
i i i i i j
ij j
ji i
i g Q Q I M F e B
dt
dB
( )wobei dBi/dt der Veränderungsrate der Biomasse von Gruppe i entspricht, gi ist die Wachstumseffizienz der Gruppe i (Produktion/Konsumption-Rate), Ii ist die Immigrationsrate, Mi ist nicht-räuberbedingte natürliche Mortalitätsrate, Fi ist die Fischerei/ Jagd- Mortalitätsrate, ei ist die Emigrationsrate und Qij ist der Anteil der Beute i im Nahrungsspektrum von Räuber j.
Vulnerabilitätswerte
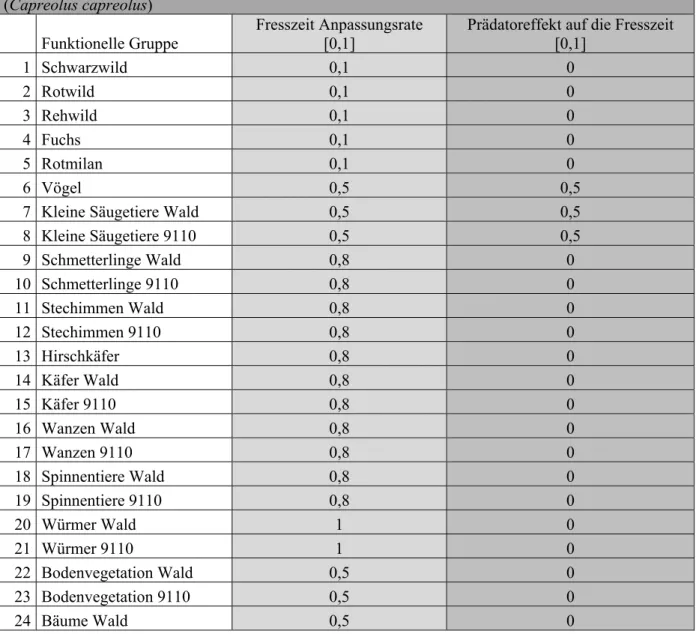
Im Kalibrierungsprozess werden die Vulnerabilitätswerte für die Räuber-Beute-Beziehungen identifiziert. Hierbei wird der Wert mit der kleinsten Quadratsumme, also geringsten quadratischen Abweichung vom Mittelwert, für das Modell übernommen (Vulnerabilitätswerte der Kalibrierung siehe Anhang). Der Vulnerabilitätswert zeigt auf, wie gefährdet die Beute durch einen Räuber ist, wobei kleine Werte anzeigen, dass nur ein kleiner Teil der Beutebiomasse dem Räuber zur Verfügung steht (CHRISTENSEN et al. 2008).
Für jede Beutegruppe werden die Ecospace-Simulationen mit drei verschiedenen Vulnerabilitätswerten durchgeführt, um die Sensitivität bezüglich dieses Parameters zu untersuchen: 1) Standardeinstellung Wert = 2, 2) maximaler Vulnerabilitätswert der Gruppe, der jedoch mindestens den Standardwert 2 beträgt und 3) mittlerer Vulnerabilitätswert der Beutegruppe in Tabelle A3.
www.nul‐online.de 8
Tabelle A2: Vulnerabilitätswerte, die während der Kalibrierung bzw. Nachbildung der historischen Daten (Abb. 2) identifiziert wurden Die funktionellen Gruppen lauten: 1: Schwarzwild (Sus scrofa), 2: Rotwild (Cervus elaphus), 3: Rehwild (Capreolus capreolus), 4: Fuchs, 5: Rotmilan, 6: Vögel, 7: Kleine Säugetiere Wald, 8: Kleine Säugetiere 9110, 9: Schmetterlinge Wald, 10: Schmetterlinge 9110, 11: Stechimmen Wald, 12: Stechimmen 9110, 13: Hirschkäfer, 14: Käfer Wald, 15: Käfer 9110, 16: Wanzen Wald, 17: Wanzen 9110, 18: Spinnentiere Wald, 19: Spinnentiere 9110, 20: Würmer Wald, 21: Würmer 9110, 22: Bodenvegetation Wald, 23: Bodenvegetation 9110, 24: Bäume Wald, 25: Bäume 9110, 26: Acker-Fauna, 27: Acker-Ernte, 28: Totholz Wald, 29: Totholz 9110, 30: Detritus, 31: Detritus 9110 Räuber: Beute:123 4 567891011121314 1516171819202126Max. Vul. Mittlerer Vul. 5 1,39 2 2 6 1 11 2 2 7 2 1 93489 934257 8 4,5 14066686418 66862129 9 1,9 1,316211,9 382 1,69111,981,052 16217 102 233122 1,771,85 1 12222 33127 112 3022 201 1 22212 306 12 2 6922 21,99 2 12221 697 132 23222 1,962 2 221,992 324 141,8 11112081,122 22 34224621,032 1208124 152 3921022 21,99 1054,982121,98336 33655 16 1,9 8212 22 2 112 8210 172 3722 21,95 2 1,5111 375 18 2300112 2 11,63221 3001302 19 2222 2 21,961,291,991,94 2 2 20 2 1,9171,852 11 174 212 281,382 21,93 8 3 2221 1 115216111 1 221,6111 163 2321 1 125,961,821,1411 2 21,98221 6 2 24 2 1 65321,988,362 21 121,96 65357 252 1 3471,883,111,991,79 2 11,992 34733
www.nul‐online.de 9
262 1 146301,21,02 21 2 12221 1,2514613 271,91 2 1,711,921,02 1,122 2 22222 32324 28 114 2 146 29 4437 4441 30 1 3,88 3,271,65 22 1 122,344 2 31 1 1,782 1,971,85 21 2 2
www.nul‐online.de 10 Definition des Fressverhaltens
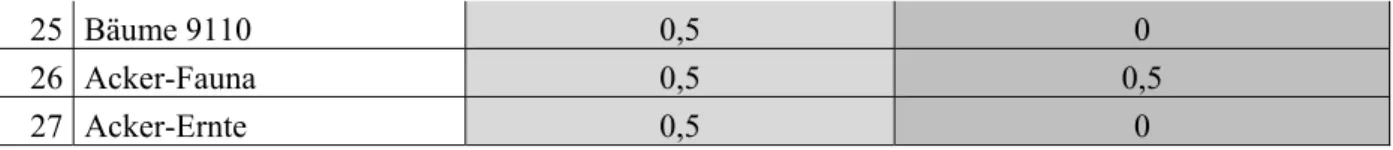
Folgende Abweichungen von den Standardeingaben wurden für das Fressverhalten der einzelnen Gruppen im Model angenommen: die Fresszeit Anpassungsrate schwankt von 0 bis 1, wobei Werte bei hohe Werte eine konstante Fresszeit andeuten, der Räuber also weder durch deine Beute oder seine Räubergruppen in der Futtersuche beeinflusst wird. Dies gilt im Modell v.a. für Würmer, die im Boden vor ihren Räubergruppen relativ sicher sind und genügend Nahrung zur Verfügung haben.
Da Räuber das Fressverhalten der Beutetiere beschränken, wird in Ecosim der Prädatoreffekt auf die Fresszeit bestimmt. Da in der Literatur keine Angaben über das Fressverhalten von Invertebraten in Anwesenheit eines Räubers gefunden wurden, wird hier davon ausgegangen, dass v.a. kleinere Säugetiere und Vögel bei Anwesenheit eines Räubers die Futteraufnahme einstellen und fliehen (Tab. A4). Die großen Säugetiergruppen und der Rotmilan haben im Modell keine Räuber zu befürchten, die ihre Fresszeit beeinflussen.
Tabelle A3: Ecosim Parameter mit Anpassungen der Fresszeit der Beutegruppen
1. Schwarzwild: Wildschwein (Sus scrofa), 2: Rotwild: Rothirsch (Cervus elaphus), 3: Rehwild: Reh (Capreolus capreolus)
Funktionelle Gruppe
Fresszeit Anpassungsrate [0,1]
Prädatoreffekt auf die Fresszeit [0,1]
1 Schwarzwild 0,1 0
2 Rotwild 0,1 0
3 Rehwild 0,1 0
4 Fuchs 0,1 0
5 Rotmilan 0,1 0
6 Vögel 0,5 0,5
7 Kleine Säugetiere Wald 0,5 0,5
8 Kleine Säugetiere 9110 0,5 0,5
9 Schmetterlinge Wald 0,8 0
10 Schmetterlinge 9110 0,8 0
11 Stechimmen Wald 0,8 0
12 Stechimmen 9110 0,8 0
13 Hirschkäfer 0,8 0
14 Käfer Wald 0,8 0
15 Käfer 9110 0,8 0
16 Wanzen Wald 0,8 0
17 Wanzen 9110 0,8 0
18 Spinnentiere Wald 0,8 0
19 Spinnentiere 9110 0,8 0
20 Würmer Wald 1 0
21 Würmer 9110 1 0
22 Bodenvegetation Wald 0,5 0
23 Bodenvegetation 9110 0,5 0
24 Bäume Wald 0,5 0
www.nul‐online.de 11
25 Bäume 9110 0,5 0
26 Acker-Fauna 0,5 0,5
27 Acker-Ernte 0,5 0
3. Ecospace
Mathematischer Hintergrund
In Ecospace wird die räumliche Verteilung der Biomasse der funktionellen Gruppe zu jeder simulierten Zeit t nach folgender Gleichung bestimmt (CHRISTENSEN et al. 2008):
i c r i c r i c r i c r i c r i c
r I gC Z E B
dt dB
, , , , , , , , ,
, ,
, ( )( )
wobei: Br,c,i beschribt die Ecopathbiomasse der Gruppe i, in einer bestimmten Zelle der Basiskarte definiert durch r: Reihe und c: Spalte der Basiskarte; Ir,c,i ist die Immigrationsrate von den umgebenden Zellen; Cr,c,i ist die Nahrungskonsumptionsrate; Zr,c,i ist die
Gesamtsterblichkeitsrate; und Er,c,i ist die gesamte Emigrationsrate von Gruppe i in der definierten Zelle.
Definition des räumlichen Verhaltens
Einige Parameter in Ecospace beschreiben das räumliche Verhalten der funktionellen Gruppen (CHRISTENSEN et al. 2008), z.B. die Dispersionsrate, die Aufschluss darüber gibt wie stark sich die Gruppen in ihrem bevorzugten und nicht bevorzugten Habitat bewegen (Tab. A5).
Tabelle A4: Ecospace-Parameter der verschiedenen Simulationen (Abb. 6, 7, Tab. 2);
die Dispersionsrate beschreibt, wie stark sich eine funktionelle Gruppe in ihrer Umgebung verbreitet bzw. bewegt
Es wurde angenommen, dass das Industriegebiet für keine Art ein bevorzugtes Habitat darstellt, bevorzugte Habitate waren 1) Hainsimsen-Buchenwald LRT 9110, 2) Wald, 3) Hirschkäferhabitate als Teil des Waldes, 4) Ackerfläche
1. Schwarzwild: Wildschwein (Sus scrofa), 2: Rotwild: Rothirsch (Cervus elaphus), 3:
Rehwild: Reh (Capreolus capreolus)
Bevorzugtes Habitat Dispersionsrate
Funktionelle Gruppe 1 2 3. 4
Basisrate (km/Jahr)
Relative Rate im nicht bevorzugten Habitat (km/Jahr)
1 Schwarzwild x x x x 30 5
2 Rotwild x x x x 30 5
3 Rehwild x x x x 30 5
4 Fuchs x x x x 30 5
5 Rotmilan x x x x 300 5
6 Vögel x x x x 300 5
7 Kleine Säugetiere Wald x x 10 2
8 Kleine Säugetiere 9110 x 10 2
9 Schmetterlinge Wald x x 3 1
www.nul‐online.de 12
10 Schmetterlinge 9110 x 3 1
11 Stechimmen Wald x x 3 1
12 Stechimmen 9110 x 3 1
13 Hirschkäfer x 3 1
14 Käfer Wald x x 3 1
15 Käfer 9110 x 3 1
16 Wanzen Wald x x 3 1
17 Wanzen 9110 x 3 1
18 Spinnentiere Wald x x 2 1
19 Spinnentiere 9110 x 2 1
20 Würmer Wald x x 1 1
21 Würmer 9110 x 1 1
22 Bodenvegetation Wald x x 0 0
23 Bodenvegetation 9110 x 0 0
24 Bäume Wald x x 0 0
25 Bäume 9110 x 0 0
26 Acker-Fauna x 5 1
27 Acker-Ernte x 0 0
28 Totholz Wald x x 0 0
29 Totholz 9110 x 0 0
30 Detritus x x x 0 0
31 Detritus 9110 x 0 0
4. Sensitivitätsanalyse
Es wurde eine umfangreiche Sensitivitätsanalyse durchgeführt. Bei dieser Analyse wird jeder Parameter um bis zu 50% erhöht oder verringert und dann die Reaktion der Parameter im Model wiedergegeben. Hier wurde jeder Parameter um 50% variiert. Als Antwortreaktion zeigte das Model Veränderungen in der ökotrophischen Effizienz einiger Gruppen, d.h. wie viel von der Biomasse der Gruppe im Model verbraucht wurde. So zeigte eine Veränderung der Biomasse des Fuchses (Gruppe 4) eine Veränderung der ökotrophischen Effizienz der kleinen Säugetiere im Wald (Gruppe 7) um 33.6% (Tab. A2). Das Model ist als robust zu bewerten, da eine Veränderung der Parameter nur geringfügige Veränderungen der ökotrophischen Effizienz zur Folge hatten und weder die Produktion, noch das Fressverhalten (Q/B) oder die Biomassen im Model änderten.
www.nul‐online.de 13 Tabelle A5: Die Ergebnisse der Sensitivitätsanalyse, bei der alle Parameter im Model um 50%
erhöht und verringert werden und dann die Antwortreaktion, also die prozentuale Veränderung der Parameter, im Model untersucht werden.
Die Tabelle zeigt die Ergebnisse, die bei 50% Variation der Eingabeparameter über 10% Veränderung im Model anzeigen.
Q/B-Parameter: Konsumption per Biomasse, EE-Parameter: Ökotrophische Effizienz
Die funktionellen Gruppen lauten: 1: Schwarzwild (Sus scrofa), 2: Rotwild (Cervus elaphus), 3:
Rehwild (Capreolus capreolus), 4: Fuchs, 5: Rotmilan, 6: Vögel, 7: Kleine Säugetiere Wald, 8:
Kleine Säugetiere 9110, 9: Schmetterlinge Wald, 10: Schmetterlinge 9110, 11: Stechimmen Wald, 12:
Stechimmen 9110, 13: Hirschkäfer, 14: Käfer Wald, 15: Käfer 9110, 16: Wanzen Wald, 17: Wanzen 9110, 18: Spinnentiere Wald, 19: Spinnentiere 9110, 20: Würmer Wald, 21: Würmer 9110, 22:
Bodenvegetation Wald, 23: Bodenvegetation 9110, 24: Bäume Wald, 25: Bäume 9110, 26: Acker- Fauna, 27: Acker-Ernte, 28: Totholz Wald, 29: Totholz 9110, 30: Detritus, 31: Detritus 9110
Variierte Eingabeparameter
Antwortreaktion
Prozentuale Veränderung im Model
Gruppe
Veränderter
Parameter Gruppe
Parameter mit anschließender
Veränderung -50% 50%
4 Biomasse 7 EE -33,6 33,6
4 Biomasse 8 EE -33,6 33,6
4 Q/B 7 EE -33,6 33,6
4 Q/B 8 EE -33,6 33,6
5 Biomasse 6 EE -39,1 39,1
5 Q/B 6 EE -39,1 39,1
6 Biomasse 5 EE -50 50
6 Biomasse 7 EE -10,8 10,8
6 Biomasse 8 EE -10,8 10,8
6 Q/B 5 EE -50 50
6 Q/B 7 EE -10,8 10,8
6 Q/B 8 EE -10,8 10,8
6 Q/B 19 EE -16 16
7 Biomasse 20 EE -14,7 14,7
7 Q/B 20 EE -14,7 14,7
8 Biomasse 21 EE -11,5 11,5
8 Q/B 21 EE -11,5 11,5
11 Biomasse 9 EE -19,4 19,4
11 Biomasse 13 EE -19,6 19,6
11 Biomasse 14 EE -12 12
11 Biomasse 16 EE -32,3 32,3
11 Biomasse 17 EE -14 14
11 Q/B 9 EE -19,4 19,4
11 Q/B 13 EE -19,6 19,6
11 Q/B 14 EE -12 12
11 Q/B 16 EE -32,3 32,3
11 Q/B 17 EE -14 14
12 Biomasse 10 EE -16,7 16,7
12 Biomasse 17 EE -16 16
12 Q/B 10 EE -16,7 16,7
12 Q/B 17 EE -16 16
14 Biomasse 18 EE -11 11
www.nul‐online.de 14
14 Q/B 18 EE -11 11
18 Biomasse 9 EE -13,2 13,2
18 Biomasse 14 EE -20,2 20,2
18 Biomasse 20 EE -32,2 32,2
18 Q/B 9 EE -13,2 13,2
18 Q/B 14 EE -20,2 20,2
18 Q/B 18 EE -23,8 23,8
18 Q/B 20 EE -32,2 32,2
19 Biomasse 10 EE -11,3 11,3
19 Biomasse 15 EE -19,4 19,4
19 Biomasse 21 EE -24,4 24,4
19 Q/B 10 EE -11,3 11,3
19 Q/B 15 EE -19,4 19,4
19 Q/B 21 EE -24,4 24,4
5. Abbildungen in Farbe
Abbildung 4: Ecospace-Basiskarte des Untersuchungsgebiets mit den Habitaten Wald (grün gemustert), Hainsimsen-Buchenwald (grünes Gebiet links in der Mitte), Industriegebiet (grau), Ackerfläche (braune Fläche) und den zwei Teilhabitaten des Hirschkäfers (rote Flächen)
www.nul‐online.de 15
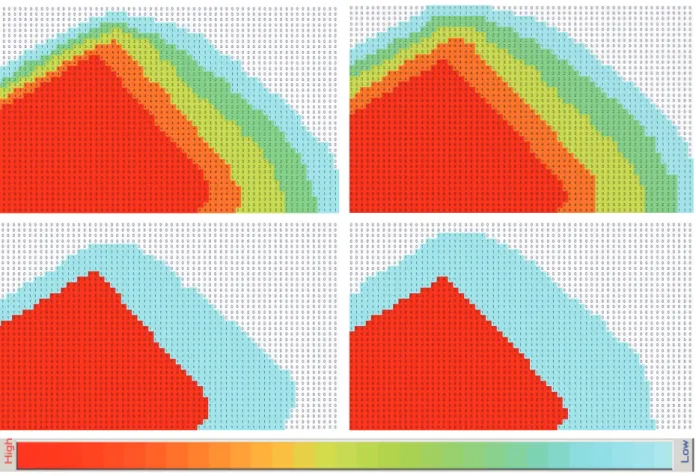
Abb. 5: Einflussbereich der Wirkfaktoren Licht (links oben) und Lärm (rechts oben), sowie Maßnahmen zur Schadensbegrenzung (unten), z.B. durch Schutzmauern. Die Intensität der Wirkfaktoren reicht in fünf Stufen von hoch bis schwach. Es wird davon ausgegangen, dass das Waldgebiet die Wirkfaktoren leicht abschirmt und dass die Feldfläche räumlich stärker beeinflusst wird.
www.nul‐online.de 16
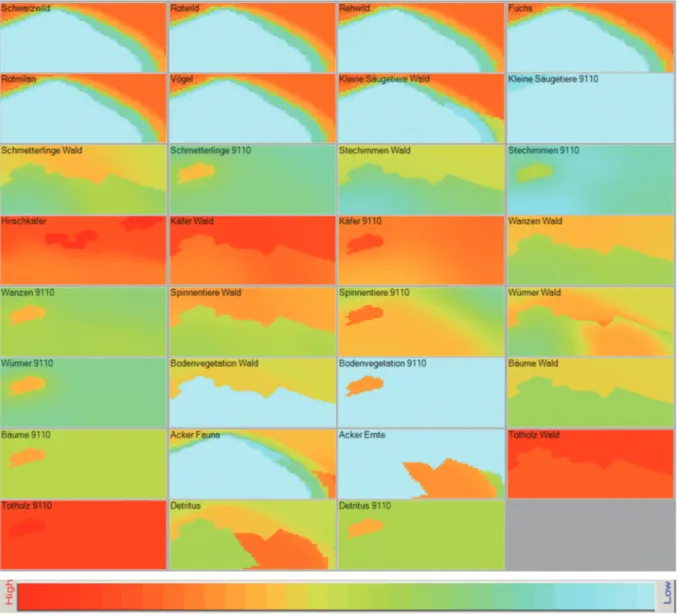
Abbildung 6: Räumliche Darstellung der Ecospace-Simulation des Habitatverlustes und der Auswirkungen der Wirkfaktoren Lärm und Licht (Abb. 5) durch den Bau des Industriegebiets (Abb. 4) für alle funktionellen Gruppen im Model (Biomassen: hoch – rot, niedrig: blau)
www.nul‐online.de 17 Abb. 7 Räumliche Darstellung der Ecospace- Simulation des Habitatverlustes und der Effektivität der Mauer zur Schadensminimierung der Auswirkungen von Lärm und Licht (Abb. 5) durch den Bau des Industriegebiets (Abb. 4) für alle funktionellen Gruppen im Modell (Biomassen: hoch – rot, niedrig:
blau)
www.nul‐online.de 18 Referenzen:
ALATALO,R.V. (1978). Bird community energetics in a boreal coniferous forest. Holarctic Ecology. 1 (4). pp. 367–376.
ALDER,S.&MARSDEN,S. (2010). Characteristics of feeding-site selection by breeding Green Woodpeckers Picus viridis in a UK agricultural landscape. Bird Study. 57 (1). pp. 100–107.
ANONYMOUS (2013). Ernte 2013: Mengen und Preise. Bundesministerium für Ernährung, Landwirtschaft und Verbraucherschutz. p. 24pp.
BADYAEV,A.V. (1997). Avian life history variation along altitudinal gradients: an example with cardueline finches. Oecologia. 111. pp. 365–374.
BANSE,K.&MOSHER,S. (1980). Adult body mass and annual production/biomass relationships of field populations. Ecological Monographs. 50 (3). pp. 355–379.
BARANCEKOVA,M. (2004). The roe deer diet: is floodplain forest optimal habitat? Folia Zoologica. 53 (3). pp. 285–292.
BARNA,M. (2001). Production of biomass of beech (Fagus sylvatica L.) leaves and buds after cutting of various intensity. Journal of Forest Science. 47 (4). pp. 164–173.
BECH,C.&PRAESTENG,K.E. (2004). Thermoregulatory use of heat increment of feeding in the tawny owl (Strix aluco). Journal of Thermal Biology. 29. pp. 649–654.
BENTON,T.G.,BRYANT,D.M.,COLE,L.&CRICK,H.Q.P. (2002). Linking agricultural practice to insect and bird populations: a historical study over three decades. Journal of Applied
Ecology. 39. pp. 673–687.
BERTOLINO,S.,VIZZINI,A.,WAUTERS,L.A.&TOSI,G. (2004). Consumption of hypogeous and epigeous fungi by red squirrel (Sciurus vulgaris) in subalpine conifer forest. Forest Ecology and Management. 202. pp. 227–233.
BIBBY,C.J.&GREEN,R.E. (1980). Foraging behaviour of migrant pied Flycatchers, Ficedula hypoleuca, on temporary territories. Journal of Animal Ecology. 49 (2). pp. 507–521.
BIEBER,C.&RUF,T. (2005). Population dynamics in wild boar Sus scrofa: ecology, elasticity of growth rate and implications for the management of pulsed resource consumers. Journal of Applied Ecology. 42. pp. 1203–1213.
BILDE,T.&TOFT,S. (2000). Evaluation of prey for the spider Dicymbium brevisetosum locket (Araneae: Linyphiidae) in single-species and mixed-species diets. Ekologia (Bratislava). 19 (3). pp. 9–18.
BLANCO,J.C.,HIRALDO,F.&HEREDIA,B. (1990). Variations in the diet and foraging behaviour of a wintering red kite (Milvus milvus) population in response to changes in food availability. Ardeola. 37 (2). pp. 267–278.
BORATYNSKI,Z.&KOTEJA,P. (2010). Sexual and natural selection on body mass and metabolic rates in freeliving bank voles. Functional Ecology. 24. pp. 1252–1261.
www.nul‐online.de 19 BOTKIN,D.B.&MILLER,R.S. (1974). Mortality rates and survival of birds. The American Naturalist. 108 (960). pp. 181–192.
BROWN,V.K. (1987). The trophic impact of small mammals in successional grasslands.
Biological Journal of the Linnean Society. 31. pp. 273–290.
CARRASCAL,L.M.,SENAR,J.C.,MOZETICH,I.,URIBE,F.&DOMENECH,J. (1998). Interactions among environmental stress, body condition, nutritional status, and dominance in Great Tits.
The Auk. 115 (3). pp. 727–738.
CAVALLINI,P. (1995). Variation in the body size of the red fox. Annales Zoologici Fennici.
32. pp. 421–427.
CHEN,B.&WISE,D.H. (1999). Bottom-up limitation of predaceous arthropods in a detritus- based terrestrial food web. Ecology. 80 (3). pp. 761–772.
CHITTY,D. (1952). Mortality among voles (Microtus agrestis) at Lake Vyrnwy,
Montgomeryshire in 1936-9. Philosophical Transactions of the Royal Society of London.
Series B, Biological Sciences. 236 (638). pp. 505–552.
CHRISTENSEN,V.,WALTERS,C.J.,PAULY,D.&FORREST,R. (2008). Ecopath with Ecosim version 6 - User Guide, Lenfest Ocean Futures Project, University of British Columbia.
www.ecopath.org.
CHURCHFIELD,S. (2002). Why are shrews so small? The costs and benefits of small size in northern temperate Sorex species in the context of foraging habits and prey supply. Acta Theriologica. 47. pp. 169–184.
CLAPHAM,S.J. (2011). The Abundance and diversity of small mammals and birds in mature crops of the perennial grasses Miscanthus x Gigantheus and Phalaris arundinacea grown for biomass energy. PhD thesis, Cardiff University .
CLAYTON,N.S.,MELLOR,R.&JACKSON,A. (1996). Seasonal patterns of food storing in the Jay Garrulus glandarius. Ibis. 138. pp. 250–255.
COHEN,A.C. (1985). Simple method for rearing the insect predator Geocoris punctipes (Heteroptera: Lygaeidae) on a meat diet. Journal of Economic Entomology. 78. pp. 1173–
1175.
CROW,T.R. (1978). Biomass and production in three contiguous forests in Northern Wisconsin. Ecology. 59 (2). pp. 265–273.
CURRY,J.P.&SCHMIDT,O.(2007). The feeding ecology of earthworms - A review.
Pedobiologia. 50. pp. 463–477.
DAAN,S.&SLOPSEMA,S. (1978). Short-term rhythms in foraging behaviour of the common vole, Microtus arvalis. Journal of comparative Physiology. 127. pp. 215–227.
DENNIS,P.,SKARTVEIT,J.,MCCRACKEN,D.I.,PAKEMANN,R.J.,BEATON,K.,KUNAVER,A.&
EVANS,D.M. (2008). The effects of livestock grazing on foliar arthropods associated with