ÄHNLICHE DOKUMENTE
A G2G analysis for the quickly evolving human immunodeficiency virus (HIV) identified strong associations of single nucleotide polymorphisms (SNPs) in the HLA class I region
The small but significant microsatellite band differentiation among natural sites and agriculturally used parcels suggests that human land use caused not only a divergence in
1) Bright day ET is expected not to change with elevation because the reduced atmospheric demand should be compensated for by a steeper leaf to air vapour pressure gradient
Field measurements of annual hor- izontal growth in the long-lived clonal plant species Carex curvula, Dryas octopetala, Salix herbacea, Vaccinium uliginosum and Empetrum nigrum
When different individuals from the same mouse line were compared, varying total numbers of Lsi1 or Lsi2 mGFP-positive GCs (or pyramidal neurons) did not affect the fractions
Of the total 450 offsprings, only 73 individuals flowered in 2004, of which 41 plants were used in the pollination experiment (3-5 plants per seed family stemming from 12
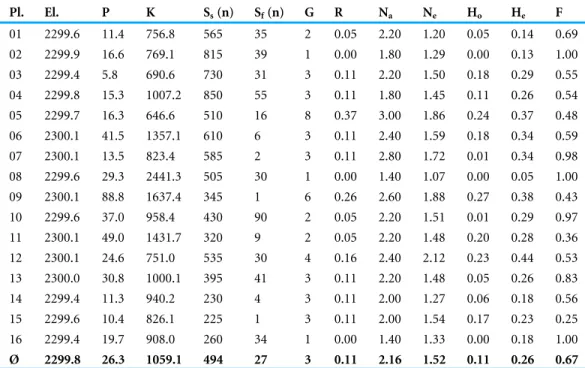
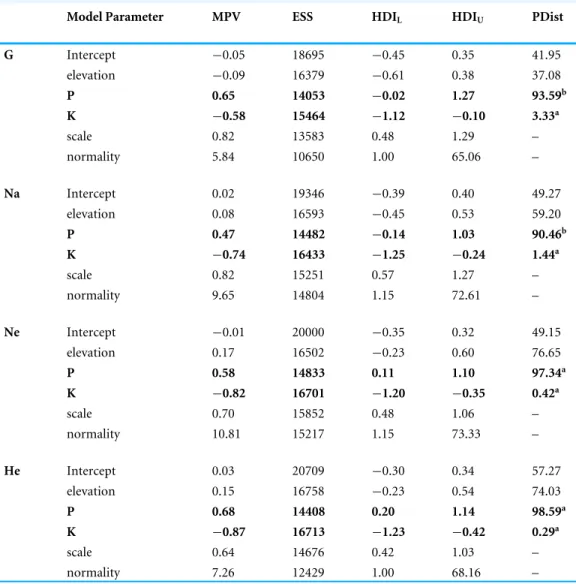
With RAPD-PCR we found a relatively high molecular diversity in the eleven populations of Scabiosa columbaria from grassland remnants in the Swiss Jura,
fimbriatus by its larger size (snout-vent length up to 200 mm vs. 295 mm), hemipenis morphology, colouration of iris, head and back, and strong genetic differentiation (4.8 %