How is the age of an anthropogenic habitat - calcareous grasslands - affecting the occurrence of plant species and vegetation composition - a historical, vegetation
122
0
0
Volltext
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(18)
(19)
(20)
(21)
(22)
(23)
(24)
(25)
(26)
(27)
(28)
Abbildung

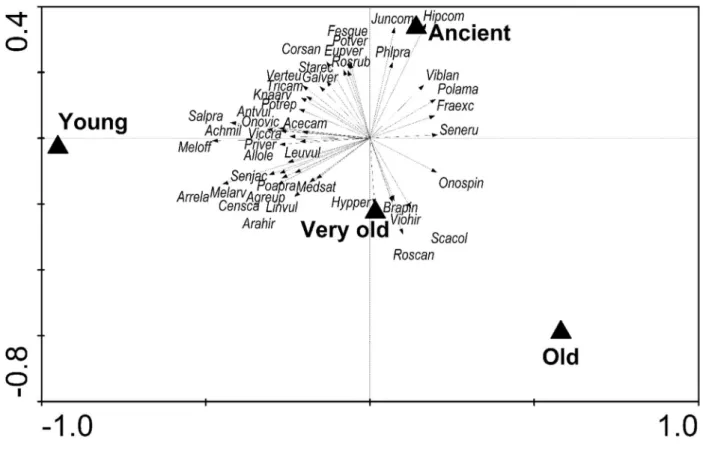
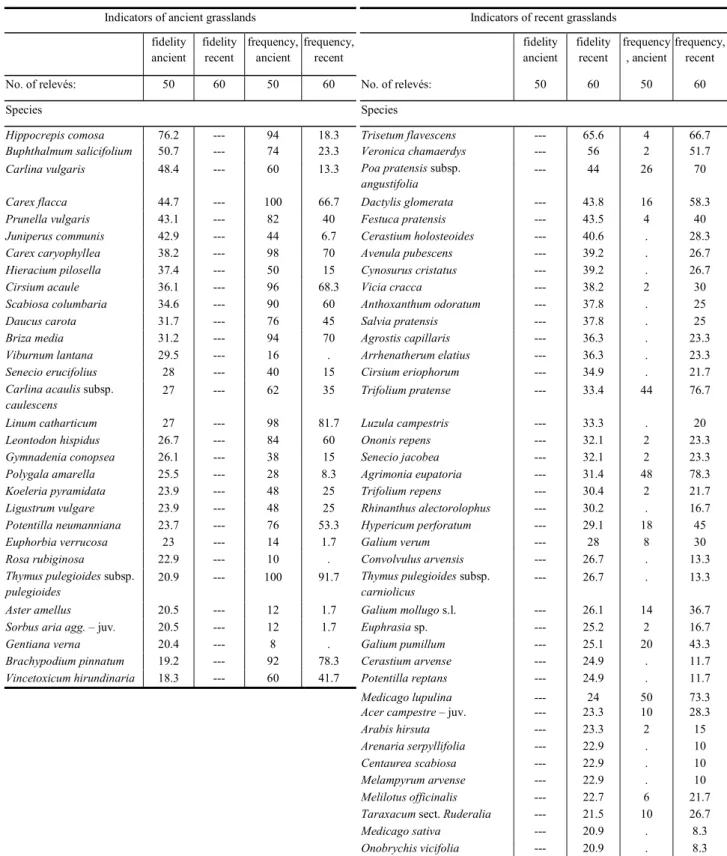
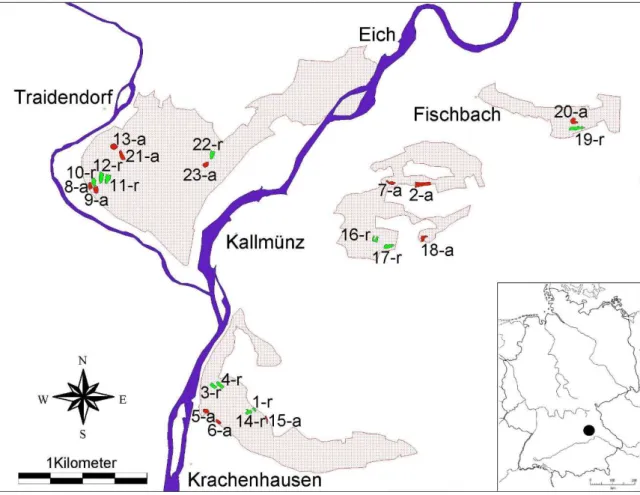
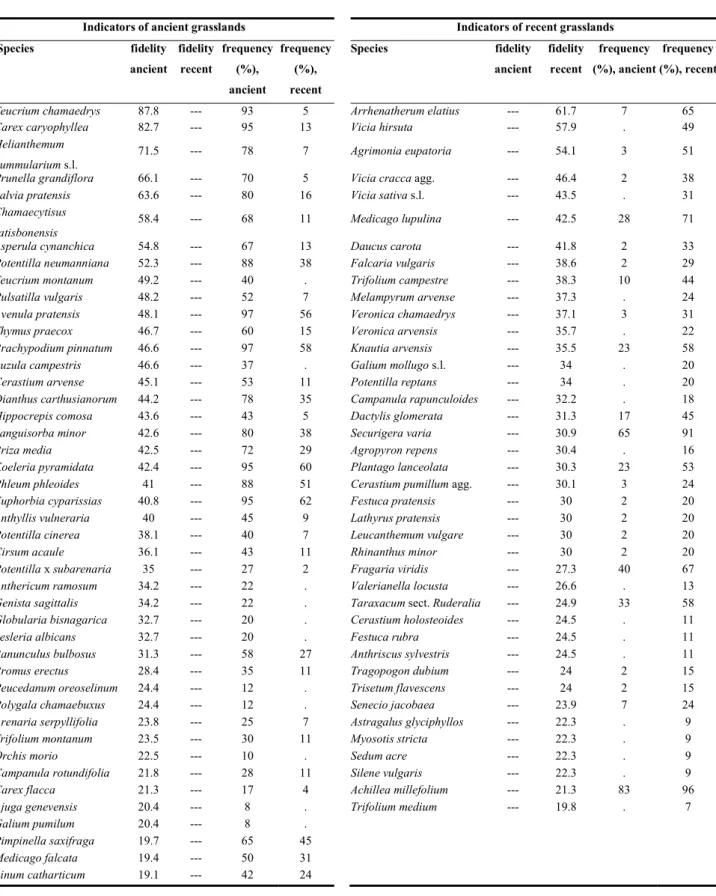
+7
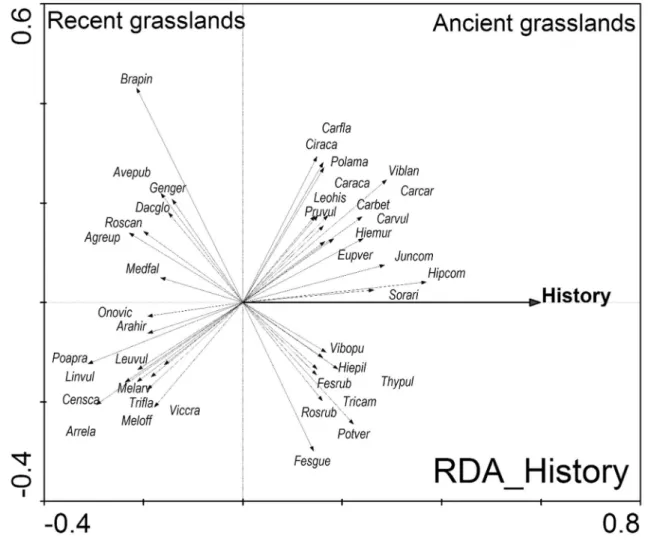
![Fig. 2.2. – Principal components analysis (PCA) presenting the correlations between a larger amount of environmental variables (geology [joMu, ox 1, ox2, ki1, qu; see Table 2.1 for details], history, altitude, inclination, PDSI of all seven months, soil](https://thumb-eu.123doks.com/thumbv2/1library_info/3849410.1515366/21.892.127.794.81.763/principal-components-analysis-presenting-correlations-environmental-variables-inclination.webp)
ÄHNLICHE DOKUMENTE
Alliaria petiolata Lasioglossum calceatum Anemone sylvestris Bombus pratorum Anemone sylvestris Bombus terrestris Anthyllis vulneraria Bombus hortorum Aquilegia vulgaris
The fourth chapter examines the effects of landscape composition, habitat connectivity and local management (grazing, mowing or abandonment) on the species
vesca (per plant) is characterized in Tab. Competition reduced shoot, root and total biomass by approximately 70% and significantly increased shoot to root
Summary of linear mixed-effect models with meadow fitted as random factor, testing the effects fertilizer (kg N ha − 1 year − 1 ; including quadratic term when significant)
To examine the effect of different types of forest edge on the specialized open- land gastropod community of nutrient-poor grasslands, we assessed the diversity and abundance of
Because unfertilized meadows harbor the most species-rich plant communities (K. Stöcklin, unpublished data), and because the higher land use diversity in Romanic
The research questions addressed in this paper are whether historical and recent observations of upper limits of vascular plants differ in altitude and if such shifts are
The regional species pool of the boreo-nemoral grasslands contains more than 400 vascular plant species, while the size of the community pool is, on average, 115–130 species;
