The role of Arabidopsis transcription factors WRKY18 and WRKY40 in plant immunity
135
0
0
Volltext
(2)(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(18)
(20)
(21)
(23)
(24)
(25)
(26)
(27)
(28)
(29)
(31)
(32)
(33)
(34)
Abbildung
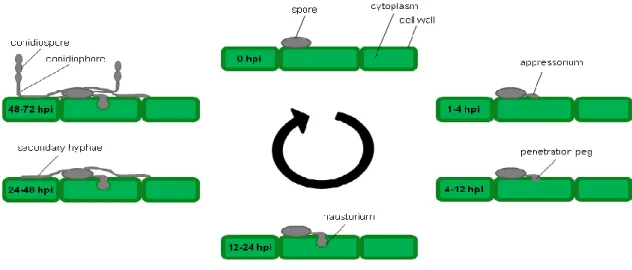
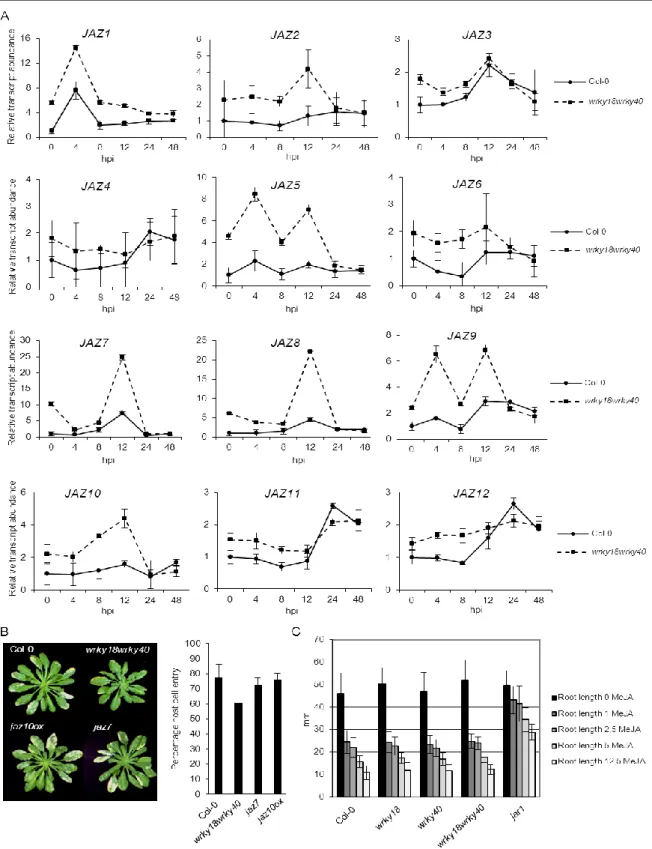
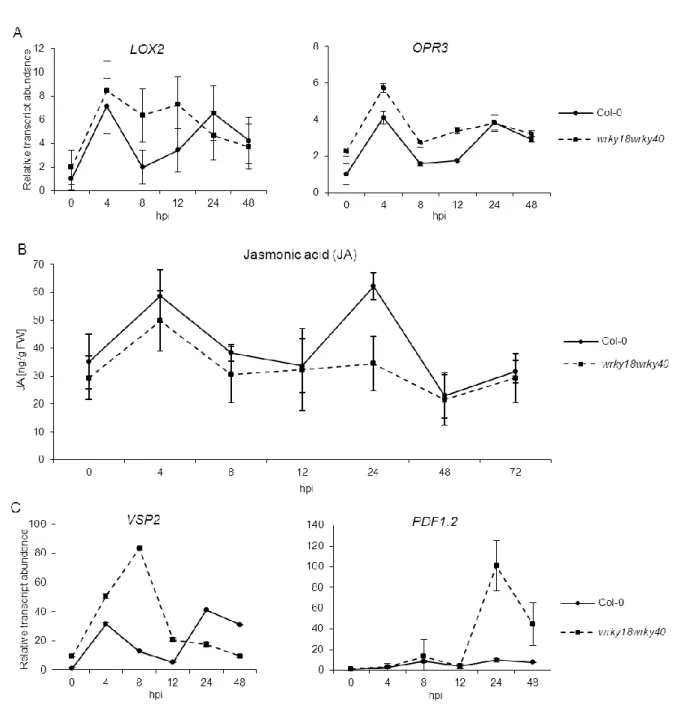
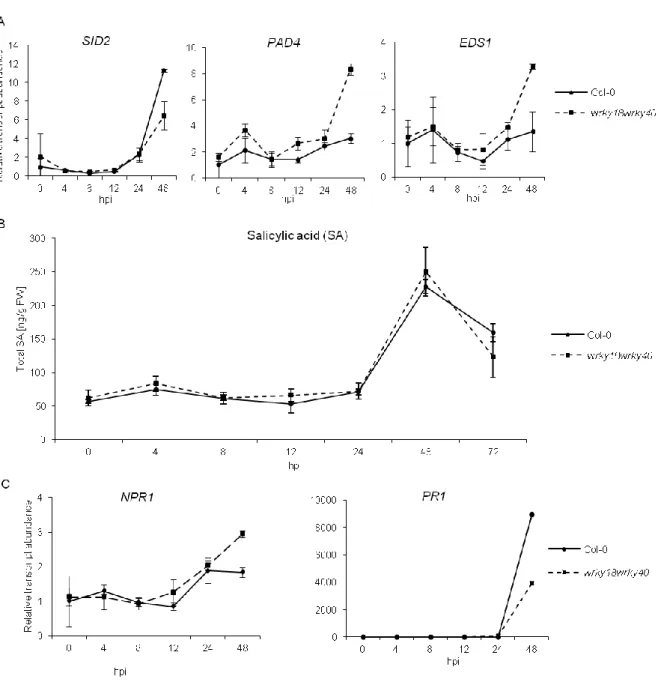
+7


ÄHNLICHE DOKUMENTE
In order to determine if PP2-B13, ACLP1, SERP1 and GRP89 proteins have roles in proper perception of other elicitors such as AtPeps and elf18 proper defense responses, we recommend
Phosphorylation-dependent differential regulation of plant growth, cell death, and innate immunity by the regulatory receptor-like kinase BAK1. Segonzac, C., and
To gain insight into the function of this gene family in dicots we expressed the BnET gene both in tobacco and Arabidopsis under the control of the viral 35S gene promoter.. This
Ever since SRF was found to regulate diverse and sometimes opposing sets of genes, ranging from genes controlling IEG response and cellular proliferation to cardiac or smooth
The differentially expressed genes in pip2;1 pip2;2 as compared to the wild type under water stresses indicate that regulation of osmotic potential and cell wall modification may
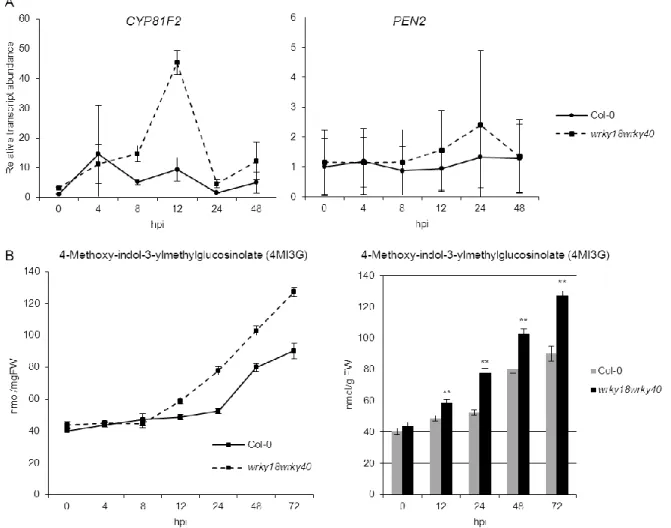
The highly correlated co-expression of MYB28/HAG1, MYB76/HAG2 and MYB29/HAG3 with aliphatic glucosinolate pathway genes (BCAT4, MAM1, CYP79F1 etc.) and phylogenetic association with
To study the function of MIKC* genes and their possible role in land plant gametophyte development and its evolution, they were isolated from a broad variety of vascular land
Specifically, GNC and GNL are able to regulate the greening in Arabidopsis through the (1) control of genes encoding for enzymes in the chlorophyll pathway (GUN5, GUN4,
