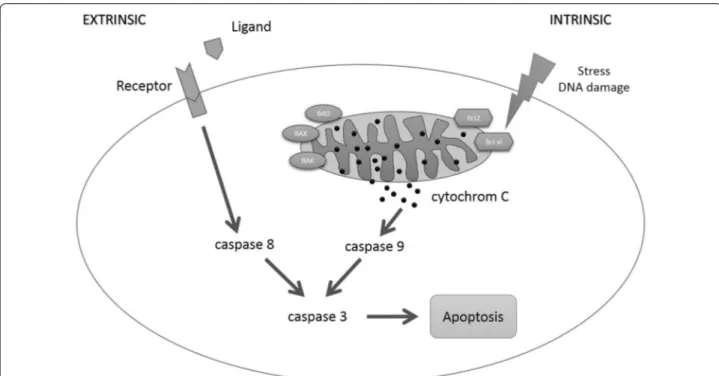
Staurosporine induces apoptosis in pancreatic carcinoma cells PaTu 8988t and Panc-1 via the intrinsic signaling pathway
8
0
0
Volltext
(2)
(3)
(4)
(5)
(6)
(7)
(8)
Abbildung
ÄHNLICHE DOKUMENTE
Expression and clinical significance of cancer stem cell markers CD24, CD44, and CD133 in pancreatic ductal adenocarcinoma and chronic pancreatitis.. Chemokine networks and
Although the use of preoperative biliary drainage is a controversy, we routinely use endoscopic stent placement in patients with cancer of the pancreatic head or extrahepatic bile
A number of research groups have shown that some established cancer cell lines, which have been maintained in culture for decades, such as head and neck squamous carcinoma
While LATS2 overexpression induces apoptosis, transverse aortic constriction (TAC)-induced cell apoptosis is inhibited by dominant negative LATS2 (DN-LATS2) over-expression
However, despite the obvious relevance of a proper regulation of adhesion molecules for the retention of circulating precursor cells to target tissues, a role of
high Plk3 detection (WS > 6) and high pT273 caspase-8 Levels (> median) in pretreatment biopsies of patients with anal carcinoma treated with definitive CRT... Figure
CTSB Deletion Reduces Apoptosis in Two Different Models of Acute Pancreatitis— To confirm that the cathepsin- and tryp- sin-mediated caspase 3 activation is translated into
A) rtTA-Ela1/tet-HA-MMP3/tet-KRas mice were treated with Caerulein and fed without Doxycycline. Wild type mice treated without Caerulein were used as control group. B)
