Edited by:
Michael Gänzle, University of Alberta, Canada Reviewed by:
Sergio I. Martinez-Monteagudo, The Ohio State University, USA Rukma Reddy, U.S. Food and Drug Administration, USA Kazutaka Yamamoto, National Food Research Institute, National Agriculture and Food Research Organization, Japan
*Correspondence:
Rudi F. Vogel, Lehrstuhl für Technische Mikrobiologie, Technische Universität München, Gregor-Mendel-Straße 4, 85354 Freising, Germany rudi.vogel@wzw.tum.de
Specialty section:
This article was submitted to Food Microbiology, a section of the journal Frontiers in Microbiology Received: 17 April 2015 Accepted: 15 June 2015 Published: 03 July 2015 Citation:
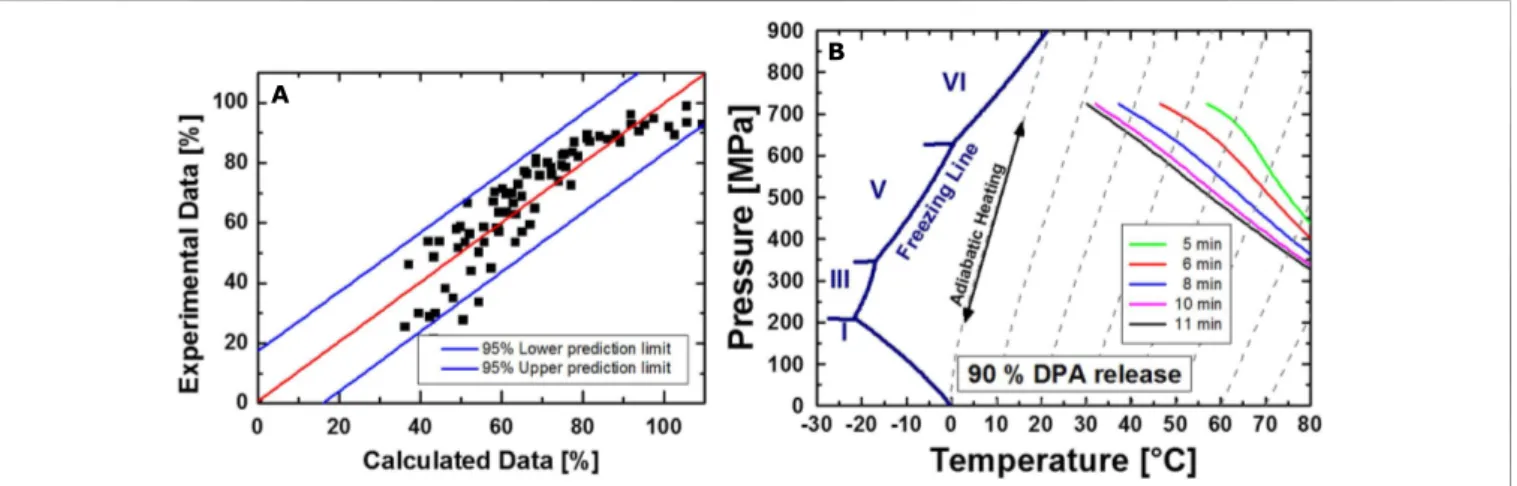
Lenz CA, Reineke K, Knorr D and Vogel RF (2015) High pressure thermal inactivation of Clostridium botulinum type E endospores – kinetic modeling and mechanistic insights.
Front. Microbiol. 6:652.
doi: 10.3389/fmicb.2015.00652
High pressure thermal inactivation of Clostridium botulinum type E
endospores – kinetic modeling and mechanistic insights
Christian A. Lenz
1, Kai Reineke
2,3, Dietrich Knorr
3and Rudi F. Vogel
1*
1
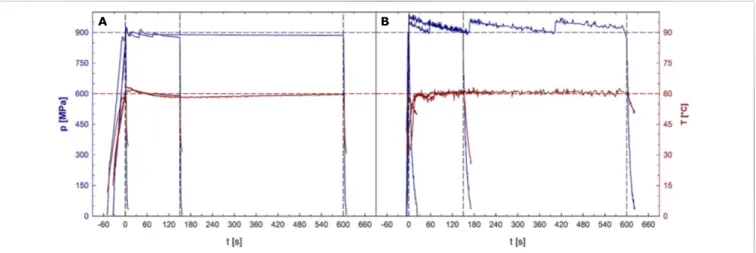
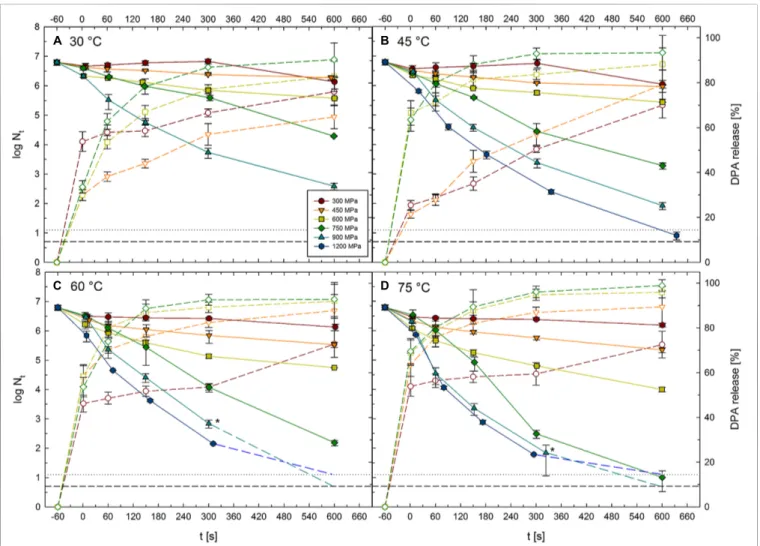
![FIGURE 2 | Mean inactivation levels of Clostridium botulinum TMW 2.990 spores after 60–600 s HPT treatments [log (N t /N 1s )] in different high pressure units](https://thumb-eu.123doks.com/thumbv2/1library_info/3882021.114327/5.892.67.827.687.977/figure-inactivation-levels-clostridium-botulinum-treatments-different-pressure.webp)
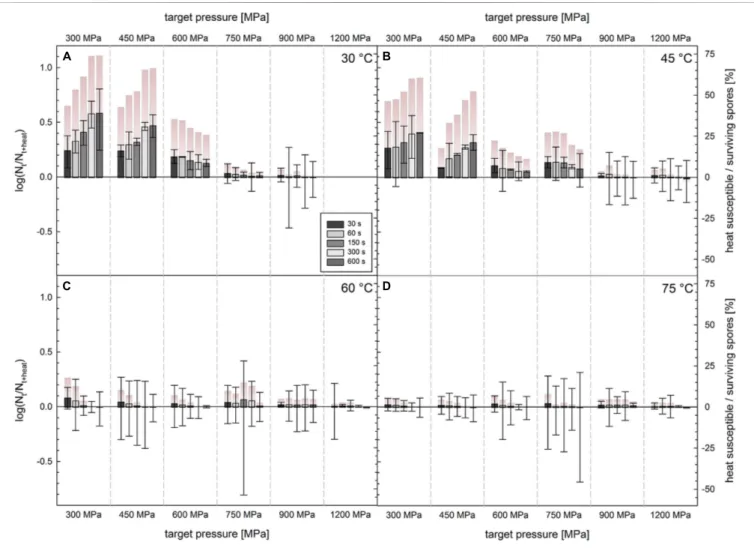
![FIGURE 5 | Difference in survivors of C. botulinum TMW 2.990 spores after HPT treatments enumerated on plate with and without lysozyme [log(N t + lysozyme /N t ); left y-axis]](https://thumb-eu.123doks.com/thumbv2/1library_info/3882021.114327/10.892.69.825.93.621/figure-difference-survivors-botulinum-treatments-enumerated-lysozyme-lysozyme.webp)