22 © F. Enke Verlag Stuttgart Zeitschrift für Soziologie, Jg. 23, Heft 1, Februar 1994, S. 22-40
Menschen, Mäuse und Fliegen
Eine wissenssoziologische Analyse der Transformation von Organismen in epistemische Objekte
Klaus Amann
Universität Bielefeld, Fakultät für Soziologie, Postfach 100131, D-33501 Bielefeld
Z usam m enfassung: Dieser Aufsatz stellt einen Beitrag zur wissenssoziologischen Analyse der epistemischen Kultur der Molekularbiologie dar. Nach einer theoretisch-konzeptuellen Verortung im Kontext der Laborstudien werden einige historische Voraussetzungen der aktuellen Entwicklungen innerhalb der Molekularbiologie skizziert.
Im Mittelpunkt des Beitrags steht die Analyse der gegenwärtigen Entwicklungen dieser Wissenschaft. Auf der Grundlage einer mehrjährigen ethnographischen Untersuchung von Wissensprozessen im naturwissenschaftlichen Labor werden deren translokale Ordnungsmuster bestimmt. Dabei wird der These nachgegangen, daß durch die Molekularbiologie eine ’neue Ordnung der (biologischen) Dinge4 (Foucault) entsteht, die durch eine Transforma
tion natürlicher Objekte in epistemische Dinge charakterisiert ist. Das Nachzeichnen dieser Entwicklung zeigt die Fruchtbarkeit des ethnographisch-mikrosoziologischen Vorgehens auch für eher traditionelle wissenschaftssoziolo
gische Fragestellungen.
1. Einleitung
Die soziologische Wissenschaftsforschung hat sich in den letzten zwanzig Jahren in weiten Teilen von einer Untersuchung der institutioneilen und nor
mativen Bedingungen von Forschung zu einer Analyse der Produktionsformen und Strukturen wissenschaftlichen Wissens gewandelt. Die empiri
schen Eigenheiten der disziplinären Gegenstände und die praktische Logik ihrer experimentellen Konstitution und Bearbeitung wurden zum Ge
genstand einer Wissenssoziologie naturwissen
schaftlichen Wissens.1
Ein konstitutives Merkmal dieses Wandels ist die Auflösung der traditionellen, in der philosophi
schen Erkenntnistheorie entwickelten Unterschei
dung eines Entdeckungs- und eines Begründungs
zusammenhangs wissenschaftlichen Wissens. Die von der funktionalistischen Wissenschaftssoziolo
gie übernommene Arbeitsteilung zwischen philo
sophischer Erkenntnistheorie auf der einen und soziologischer Analyse der institutionellen und normativen Erkenntnisbedingungen auf der ande
ren Seite wurde durch empirische Analysen histo
rischer und zeitgenössischer Wissensproduktion radikal in Frage gestellt. Neben den frühen A rbei
ten von Ludwik Fleck (1935) waren es auch die Ar
beiten von Thomas Kuhn (1962), die andere Unter- 1 Vgl. dazu Knorr-Cetina/Mulkay (1983a) Pickering (1992) und insbesondere Heintz (1993), die eine aus
führliche Darstellung der verschiedenen neueren Ansätze in der Wissenschaftsforschung gibt.
Scheidungen für die Analyse (naturw issenschaft
lichen Wissens einforderten.
D er Übergang von ’externen4 Betrachtungen der Strukturen und Institutionalisierungsformen von Wissenschaft zur Betrachtung als Tätigkeiten mit empirisch zu untersuchenden Eigenheiten fand unter dem Einfluß erkenntnistheoretischer Über
legungen statt.2 Wissenschaftshistorisch und er
kenntnistheoretisch argumentierende Relativie
rungen einer aus den wissenschaftlichen Begrün
dungszusammenhängen kausal ableitbaren ’Logik der Forschung4, der erkenntnistheoretische Nach
weis der Unterdeterminiertheit von wissenschaft
lichen Theorien durch Beobachtungen bzw. der Theoriegeladenheit wissenschaftlicher Datenpro
duktion eröffneten u.a. die Möglichkeit, naturwis
senschaftliche Wissensproduktion als eine an loka
le Kontexte gebundene und soziale Tätigkeit zu betrachten und diese soziologische black box mit empirischen Mitteln zu durchleuchten.3
Wenn empirische Beobachtungen - wissenschaft
liche Tatsachen - in unterschiedlichen Theorien (Erklärungsmodellen) unterschiedliche, aber theoriebezogen konsistente Bedeutungen erlan
gen können und umgekehrt formulierte wissen
schaftliche Tatsachen stets als Resultat theorieab
hängiger Beobachtungen und nicht mehr in einem
2 In diesem Zusammenhang sind u. a. die Arbeiten von Quine (1953), Feyerabend (1975) und Hesse (1980) relevant.
3 Vgl. dazu Knorr-Cetina/Mulkay (1983b),
Klaus Amann: Menschen, Mäuse und Fliegen 23 naiven Sinne als ’Deskriptionen4 gesehen werden
können, ihr Gegenstand somit nicht mehr als E nt
deckung eines objektiv Vorhandenen zu bestim
men ist, dann werden verschiedene praktische Konsequenzen für die Untersuchung und Darstel
lung moderner Wissenschaften möglich.
U.a. wurden folgende Strategien daraus abgelei
tet: aus wissenschaftshistorischer Perspektive eine Strategie, die rekonstruktiv Abfolgen von Erklä
rungsmodellen, Theorien und damit verbundenen Formen der wissenschaftlichen Praxis sowie die Untersuchung von Übergangsphasen zum Gegen
stand machen kann; aus wissenschaftshistorischer wie -soziologischer Perspektive die systematische Suche und Analyse ’offener’ Situationen im For
schungsprozeß, nämlich dort wo eine Differenz zwischen Interpretationen von wissenschaftlichen Beobachtungen expliziert und als Kontroverse ausgetragen wird (MacKenzie 1981; Pickering 1984; oder auch Collins 1985); aus wissenssoziolo
gischer Perspektive die mikroskopische Analyse naturwissenschaftlichen Wissens.4
Gemeinsam ist vielen der jüngeren Arbeiten eine Hinwendung zu empirischen Bearbeitungen ihrer Fragestellungen, die sich nicht allein auf die Ex
ploration der Darstellungsebene der Wissenspro
duzenten beschränken, sondern in die Mikropro
zesse der jeweiligen Lebenswelten Einblick zu nehmen versuchen.5
Zu letzteren gehören insbesondere die Arbeiten von Latour und Woolgar (1979), Lynch (1985) und Knorr-Cetina (1981), die die wissenschaftliche Ar
beit in Forschungslaboren in den M ittelpunkt der Analyse stellen. Die Konzeption vom Labor als Ort und sozialer Form der Wissensproduktion wird von diesen Autoren in unterschiedlicher Weise weiterentwickelt. Bei Latour (1987), Law und Cal- lon (1988) werden die Verknüpfungen zwischen Akteuren und ’natürlichen4 Gegenständen, zwi
schen Macht und Wissen als zu sozialen Beziehun
gen gewebtes und mit rhetorischen und (macht)politischen Mitteln verwobenes Netzwerk konzipiert und exemplarisch rekonstruiert. ’Tech-
4 Bloor (1976) bestimmt diese Wissenssoziologie zu
erst als ein ’strong programme4, das dann durch das sog. Interessenmodell (Bames/MacKenzie 1979) in
terpretiert wird. Vgl. dazu auch Heintz (1993):
535 ff. Andere Arbeiten wie die von Latour/Woolgar (1979), Lynch (19^5) und Knorr-Cetina (1981) folgen diesem Ansatz nur bedingt.
5 Oder sie tun dies in einer spezifischen Weise, indem sie Darstellungspraktiken diskursanalytisch behan
deln (z.B . Mulkay et al. 1983).
nische4 und ’soziale4 Beziehungen werden auf epi- stemologischer Ebene ebensowenig unterschieden wie menschliche und nicht-menschliche Akteure im Sinne semiotischer Konzepte egalisiert werden.
Lynch und andere ethnomethodologische Autoren wie Livingston (1986) oder Garfinkei (Garfinkei et al. 1981) verstehen ihre Beiträge als Teil der
’ethnomethodological studies of work4, in denen sie die besondere Form wissenschaftlicher Labor
arbeit zu ihrem Gegenstand machen.6
Knorr-Cetina schließlich erweiterte den Laborstu
dienansatz zu einer kulturanalytischen Konzep
tion, die im Folgenden Hintergrund meiner Dar
stellung sein wird. Sie entwickelte auf der Grund
lage von Laborstudien in der Elementarteilchen
physik und der Molekularbiologie (z.B. Knorr- Cetina 1992b, Amann/Knorr-Cetina 1988) das Konzept epistemischer Kulturen (1994). Vorrangig geht es dabei um die Systematik der unterscheid
baren kulturellen Ordnungen, mit denen etwa in biologischen oder physikalischen Kontexten Wis
sen erzeugt wird. Sichtbar gemacht wird die ’dis
unity of the sciences4 (Knorr-Cetina) nicht nur als eine Oberflächendifferenz ihrer Forschungspraxis, sondern als Unterschiedlichkeit ihrer Erkenntnis
kulturen. D er empirische Vergleich von For
schungsfeldern wird fruchtbar gemacht um diese systematischen Differenzen herauszuarbeiten.7 Die Mikroanalysen wissenschaftlicher Laborarbeit handelten sich in unterschiedlicher Weise den Vor
wurf ein, zugunsten der Detaillierung und Präzi
sierung lokaler Ereignisse die translokalen, univer
sellen Ordnungsmuster von Wissenschaft, The
men, die bislang in der Wissenschaftsforschung be
arbeitet wurden, aus dem Blick zu verlieren.8 Hin
zu kommen kritische Auseinandersetzungen mit den unterschiedlichen Konzeptionen von Kon
struktivismus, die (nicht nur) in der neueren Wis
senschaftsforschung und -theorie Verwendung fin
den.9
Nicht nur aus dieser Kritik wird deutlich, daß sich neben den - hier nicht weiter explizierten - Diffe
renzen zwischen den aufgeführten Autoren und Arbeiten eine dem wissenssoziologischen Vörge- 6 Vgl. dazu auch die Darstellung von Lynch (1992).
7 Dieses Vorgehen wird in Knorr-Cetina (1992b) auf weitere Bereiche ausgedehnt. Vgl. auch Knorr-Ceti
na/Amann (1992).
8 Vgl. dazu etwa Nowotny (1984), Cozzens/Gieryn (1990).
9 So z.B . zuletzt Sismondo (1993). Vgl. dazu Knorr- Cetina (1993) und (1989). Darauf kann in diesem Zusammenhang nicht näher eingegangen werden.
24 Zeitschrift für Soziologie, Jg. 23, Heft 1, Februar 1994, S. 22-40 hen gegenüber indifferente oder ablehnende Wis
senschaftsforschung identifizieren läßt.
Diese eher traditionell orientierten wissenschafts
soziologische Ansätze adaptieren zwar z.T. einen erkenntnistheoretischen Relativismus etwa in Form des radikalen Konstruktivismus, halten je
doch u.a. an der kognitiven Einheit der Wissen
schaft fest und explorieren jenseits dieser Voraus
setzung die sozialen Dimensionen von Wissen
schaft. Sie interessieren sich insbesondere für Fra
gen, die eine stärker makroskopische Betrachtung von übergreifenden Entwicklungstendenzen gan
zer Wissenschaftsbereiche oder des Wissenschafts
systems schlechthin erfordern und an der Unter
scheidung von gesellschaftlichen Funktionssyste
men orientiert sind.10
Die Vermittlung zwischen diesen letztgenannten Ansätzen und der Wissenssoziologie naturwissen
schaftlichen Wissens ist von verschiedenen wech
selseitigen Miß- und Unverständnissen gekenn
zeichnet, die einer eigenen Analyse bedürften und die hier nicht versucht werden soll.11 Ich will dage
gen im Weiteren ethnographisches, mit mikroana
lytischen Verfahren gewonnenes Datenmaterial als Basis für die Interpretation translokaler Entwick
lungen in der Molekularbiologie benutzen. Dazu werde ich aus den Innenansichten lokaler For
schungsprozesse eine Systematik für deren wis
senssoziologische Rekonstruktion entwickeln. Mit dieser Rekonstruktion soll die Leerstelle zwischen einer wissenssoziologischen Entzifferung natur
wissenschaftlicher Tatsachenproduktion und einer von ’außen4, d .h . einer das Wissenschaftssystem und seine Eigenheiten beschreibenden Makroper
spektive gefüllt werden. Ich werden mich dazu ei
ner kulturalistischen Perspektive bedienen. In de
ren M ittelpunkt rücken die besonderen Objekte der Molekularbiologie, deren Konstitution und Transformation.
In Anlehnung an Knorr-Cetina verstehe ich als epistemische Kultur der Molekularbiologie dieje
10 Um nur exemplarische Beispiele zu nennen: Hohn/
Schimank (1990), Nelkin (1987) oder Luhmann (1990).
11 Einen Beitrag zu dieser Debatte liefern Hasse et al.
(1993). Dies bedeutet nicht, daß gerade an den Stel
len, an denen es um die empirische Analyse konkre
ter Wissenschaftsfelder geht, keine konsens- oder an
schlußfähigen Resultate vorliegen. (Vgl. dazu etwa Bonß et al. 1993). Entsprechendes kann man u.U.
auch von einem Vergleich der noch nicht publizierten Resultate aus dem von Zimmerli geleiteten Projekt zur Technologisierung der Biologie und den in die
sem Aufsatz diskutierten Phänomenen erwarten.
nigen Einheiten wissenschaftlicher Praxis, die - empirisch nachvollziehbar - zusammenhängende Formen, Strukturen und Mechanismen der Wis
senserzeugung, Stabilisierung und Diffusion bil
den. D er praktische Zugang zu diesen Einheiten erfolgt über eine Exploration der Arbeitsplätze, der O rte, an denen die Gegenstände und Themen wissenschaftlichen Wissens in materialer und sym
bolischer Weise bearbeitet werden. Diese Arbeits
plätze sind in den experimentellen Wissenschaften Laboratorien.
Über solche Zugänge und mehrjährige ethnogra
phische Beobachtungen in molekularbiologischen Forschungslabors in Deutschland ist die empiri
sche Grundlage dieses Aufsatzes entstanden. Ne
ben einer detaillierten Untersuchung der prakti
schen Strukturen von Wissensprozessen (Amann 1990) ermöglichten diese Beobachtungen einen Einblick in übergreifende Entwicklungen inner
halb der modernen (Molekular-)Biologie. Mein Ziel ist, einen zentralen Aspekt ihrer epistemi- schen Kultur in den Mittelpunkt der folgenden Ar
gumentation zu rücken: die materiale und symboli
sche Transformation *natürlicher4 Objekte in episte
mische Dinge.
Naturwissenschaftliches Experimentieren und Wissen ist für uns in selbstverständlicher Weise mit der Vorstellung verbunden, daß das erstere an der Gegenständlichkeit der Welt seinen Ausgang nimmt und das letztere in irgendeiner systemati
schen Weise ein Resultat dieses gegenstandsorien
tierten Tuns ist. Für den soziologischen Beobach
ter im molekularbiologischen Forschungslabor ist die Vielfalt und Massivität solchen Tuns nicht nur ein Grund für anfängliche Verstehensprobleme ge
genüber dem konkreten Forschungsalltag. Sie be
deutet auch eine konzeptuelle Herausforderung, eine feldadäquate Rekonstruktion der Einheiten der beobachteten Wissensprozesse zu entwickeln.
Gegenüber einer Trennung in Elemente der Theo
rie, handelnde Akteure, technische Innovationen und Gegenstände wissenschaftlichen Wissens, die jeweils für sich genommen zum Thema gemacht werden könnten, legt eine empirische Analyse der entsprechenden disziplinären Praktiken die wis
senssoziologische Rekonstruktion experimenteller Modellsysteme als zentrale Einheiten molekular
biologischer Wissensprozesse nahe.
Experimentelle Modellsysteme werde ich als den Kern einer besonderen materialen Gestalt und Ge
staltung molekularbiologischer Forschungsgegen
stände betrachten. Bei ihrer wissenssoziologischen Analyse werden die dazugehörigen empirischen
Klaus Amann: Menschen, Mäuse und Fliegen 25 und theoretischen Gegenstände als eine neue Ord
nung der Dinge (Foucault) bestimmt. Die These dieses Aufsatzes ist folgende: Mit der Molekular
biologie befindet sich ein zentraler Bereich der modernen Biologie auf dem Weg zu einer Techno- Wissenschaft, zu einer Wissenschaft, die ihre G e
genstände - Lebewesen und Teile davon - nicht nur mit immer raffinierteren technischen Appara
turen behandelt, zerlegt, prozessiert, analysiert und verändert. Sie konstituiert vielmehr in einem elementaren und neuen Sinn bio-logische Objekte als Technofakte, die nicht mehr als Veränderungen einer wie auch immer zu bestimmenden ’natürli
chen N atur4 beschrieben werden können und mit denen zugleich die bisherige (epistemologische und disziplinäre) Ordnung der Biologie ins Wan
ken gerät.
Es geht dabei um mehr und um anderes als die populärwissenschaftlich verbreiteten angsterzeu
genden Thesen von der Technisierung des Leben
digen4 oder von der ’Verlegung der Schöpfung ins Labor4 implizieren.12 Aus der Beobachtung lokaler Forschungspraxis wird vielmehr die Etablierung einer neuen Ordnung, einer neuen Bio-Logie sichtbar. Im Mittelpunkt dieser neuen Ordnung stehen Modellsysteme, d.h. epistemische Objekte und Identitäten, die die sogenannten natürlichen Objekte - Pflanzen und H ere, Menschen, Mäuse und Fliegen - als in einer externen Natur vorfindli- che Einheiten biologischen Wissens ablösen. Die
ser Prozeß ist weder abgeschlossen, noch ist er ohne weiteres als Entwicklungslinie in die Zukunft der Disziplin prognostisch zu verlängern. Gleich
wohl können verschiedene Elemente dieser neuen Ordnung klar bestimmt werden.
2. Von den ’natürlichen A rten 6 zur gentechnischen Grenzüberschreitung Bevor ich die Argumente und empirischen A n
haltspunkte für meine These darlege, sei - in star
ker Verkürzung - das im beginnenden 19. Jahrhun
dert etablierte Bild der Wissenschaft vom Leben
digen wiedergegeben, das uns heute aus unserer Schulbildung vertraut ist.
Für den Übergang von der Naturgeschichte zur modernen Biologie als der Wissenschaft vom Le
bendigen gibt Michel Foucault in seiner ’Ordnung der Dinge4 (1971) eine auf die neue Ordnungs-Lo
gik der Biologie orientierte Darstellung. E r hebt 12 12 Formulierungen, wie man sie stellvertretend für an
dere bei Herbig (1982) findet.
dabei die neue Konzeption der Lebewesen als or
ganisierte Entitäten hervor, die nun aufgrund der unterschiedlichen Organisation ihrer elementaren Lebensfunktionen in hierarchische Klassifikatio
nen geordnet werden und aufgrund von sichtbaren und unsichtbaren Merkmalen voneinander zu un
terscheiden sind. Eine Ordnung, in der nicht mehr die Geschichte einer Pflanze oder eines Tieres ge
schrieben wird, zu der auch gehörte, „ . . . zu sa
gen, welches ihre Elemente und ihre Organe, wel
ches die Ähnlichkeiten, die man in ihnen finden kann, welches die Kräfte, die man ihnen zu
schreibt, die Legenden oder Geschichten, mit de
nen sie vermischt werden, die Wappen, auf denen sie zu sehen sind, und die Medikamente, die man aus ihrer Substanz herstellt, die Nahrungsmittel, die sie bieten, gewesen sind.44 (Foucault 1971:169) Die sezierende und mit den Mitteln der Biochemie und -Physik ausgestattete Biologie folgt - neben den Aufgaben des Sammelns, Ordnens und Be- nennens der Vielfalt des Lebendigen - den früh identifizierten Lebensfunktionen und ihren (mi
krobiologischen Realisierungsformen. Taxonomi- sche Unterscheidungen und funktionale Struktu
ren sind dabei auch Möglichkeiten der Abgren
zung, Unterscheidung und Etablierung subdiszi
plinärer Einheiten der Biologie.
Funktionale Prinzipien wie Fortpflanzung, Ver
dauung, Atmung, Körperbau sind zum einen Ähn
lichkeiten und Differenzen, die als Elemente von Taxonomienbildung fungieren. Mit diesen wird über die Individuen und A rten hinweg ein Gewebe von funktionalen Beziehungen etabliert. Zum an
deren lassen diese Taxonomien die natürlichen Unterschiede als Unterschiede wirklicher Tiere und A rten fortbestehen und ermöglichen, indivi
duelle Veränderungen als individuelle Variationen zu identifizieren. Dieses Bild einer Ordnung des Reiches des Lebendigen wird mit Darwin und der Evolutionstheorie zu einem einheitlichen, über die Zeit sich entwickelnden Stammbaum alles Leben
digen, einer natürlichen Ordnung, geformt.
Die moderne Genetik startet auf diesem Boden der natürlichen Vielfalt des Lebendigen und der Unterscheidbarkeit von A rten, baut aber auf die individuellen Variationen als M utationen, um von ihnen zur Charakterisierung der artspezifischen Normalität zu gelangen. Sie nutzt die sichtbaren Variationen natürlicher Lebewesen sowohl als ge
nealogischen Effekt - zur Untersuchung der Orga
nisation der Reproduktionsmechanismen (Men
del) - als auch als Zeichen genetischer Verände
rungen. Sie befreit sich schrittweise von der Be
26 Zeitschrift für Soziologie, Jg. 23, Heft 1, Februar 1994, S. 22-40 schränkung natürlicher Variationsbreiten der Ar
ten, zum einen durch Zuchtwahl, zum anderen durch eigene Mutationsstrategien.13
Mit der Etablierung der DNS als Baustoff und Substanz der Chromosomen, als Träger der Erbin
formation und als Bauplan des Lebendigen durch die Molekularbiologie ist ein Punkt in der Ent
wicklung der modernen Biologie erreicht, an dem die Grundfrage ’Was ist Leben?’14mit der Entziffe
rung der genetischen Schriftrolle eine mögliche Antwort finden kann. Die biochemische und bio
physikalische Bearbeitung dieser Schriftrollen zeigt die Chance einer großen neuen Vereinheitli
chung: Leben als Variation des Immergleichen von vier Aminosäuren in Form von Nukleinsäuremole
külen.
Merkmale und Funktionen, die zusammen Phäno
typen bilden, lösen sich in die einheitliche Spra
che, das einheitliche Alphabet der Gene auf. Die Strategien der Entzifferung der Texte, die mithilfe dieses Alphabets geschrieben sind, lösen den mo
lekularbiologischen Forschungsprozeß schrittwei
se von den besonderen Organismen und führen zur Akkumulation eines exponentiell wachsenden Korpus von molekulargenetischen und - biologi
schen Techniken.
In der klassischen Genetik, insbesondere bei der A rbeit mit der Fruchtfliege Drosophila melanoga- ster, wurde ein umfangreicher Wissenskorpus über den Zusammenhang von Veränderungen bestimm
ter chromosomaler Abschnitte und Veränderungen des Phänotyps erarbeitet (Kohler 1993). Geneti
sche Karten dokumentieren dieses Verhältnis von Merkmalen und genetischem/chromosomalem Ort. Weniger umfassend, aber ebenfalls intensiv sind die Arbeiten an der Maus. H ier werden syste
matisch M utationen als Ausgangspunkt für die Etablierung von Tierstämmen genutzt, die einen materiellen Fundus für Forschungsarbeiten bilden.
Aus der Virologie und Bakteriologie kommen E nt
wicklungen, die Grundlage einer Gentechnologie werden: die Herauslösung und die Vervielfältigung von DNS-Stücken unabhängig von ihrem Ur
sprungsorganismus. Erst aus der Zusammenfüh
rung von Genetik und Gentechniken ergibt sich eine neue Ebene der Beschäftigung mit dem Le
13 Kohler (1993) zeigt in seiner Geschichte der Droso- phÜa-Forschung, wie durch die Entwicklung entspre
chender Mutationsstrategien eine massive Auswei
tung der Forschung an der Fruchtfliege zu Beginn dieses Jahrhunderts erfolgen konnte.
14 Die Ausgangsfrage für Erwin Schrödinger (1949).
bendigen: Biologie als Arbeit am genetischen Mate- rial. Wo zuvor ein Vergleich auf die Merkmals- und funktionale Ebene beschränkt war, ergibt sich nun ein basales Vergleichen: die Parallelisierung von Genstrukturen verschiedenster Organismen.
Die Gentechniken überschreiten scheinbar mühe
los die innerdisziplinären Schranken der Biologie.
Mit ihnen findet eine Homogenisierung der For
schungspraxis quer zu den bisherigen Unterschei
dungen statt. Sowohl die professionsbiographische H erkunft von Biologen - und selbst das ist keine Beschränkung mehr, es gibt im molekularbiologi
schen Labor Mediziner, Chemiker und Physiker ebenso wie Tierernährungskundler - als auch der Ursprung des Untersuchungsmaterials wird irrele
vant. Es entsteht ein interdisziplinärer Diskurs entlang sich überlagernden Praktiken.
Im folgenden werde ich solche Praktiken und die Systematik der verschiedenen Grenzüberschrei
tungen auf der Grundlage eines umfangreichen ethnographischen Datenkorpus skizzieren. Dabei wird deutlich werden, daß nicht die Exegese fach
wissenschaftlicher Lektüre oder die Durchführung von Experteninterviews, sondern nur eine detail
lierte ’Vor-Ort‘ Exploration diese Systematik ent
ziffern kann.
3. M odellsystem e als Technofakte und epistem ische O bjekte
Ich werde mich in der weiteren Darstellung zwi
schen zwei Blickwinkeln auf aktuelle Entwicklun
gen in der Molekularbiologie bewegen. Auf der ei
nen Seite stehen Beobachtungen, die translokale Voraussetzungen fü r Lokales beschreiben. Auf der anderen Seite stehen Beobachtungen, die die lo
kale Organisation von Forschungs- und Wissens
prozessen beschreiben.
Dazu werde ich im folgenden Abschnitt den Über
gang von der Untersuchung von Organismen als vorgegebenen Einheiten der Natur zur Bearbei
tung von biologischem Untersuchungsmaterial darstellen. In Verbindung damit steht die Etablie
rung von komplexen analytischen Mitteln als laborinterne und externe Infrastruktur von mole
kularbiologischen Laboratorien. In einem weite
ren Abschnitt (3.2) werde ich zwei miteinander zu
sammenhängende neue Strukturelemente darstel
len, die erkennbar machen, wie die Transforma
tion natürlicher in epistemische Objekte in der m aterialen Organisation des Forschens vollzogen wird. Die Folge ist das Verschwinden von natürli
Klaus Amann: Menschen, Mäuse und Fliegen 27 chen Organismen als Lieferanten biologischen
Materials und als direkte Referenzobjekte des Wissensprozesses, wie zugleich die Entstehung neuer Einheiten. Schließlich werde ich Verknüp
fungspunkte zwischen der analytischen Infrastruk
tur und diesen Strukturelementen exemplarisch aufzeigen, an denen die neuen biologischen Ob
jekte Grundlagen für die Darstellung neuen Wis
sens liefern. (3.3)
3.1 Biologisches Material und analytische Infrastruktur
Vor der Etablierung von biologischen Technofak
ten stand die Entwicklung von vielfältigen Verfah
ren der Umwandlung lebendiger Organismen in biologisches Material. Diese Umwandlung setzt zweierlei voraus. Zum einen erfordert sie eine tay- loristische Betrachtung von Organismen: Organis
men können danach (auch) als primäre Produk
tionszusammenhänge von Organen, Zellen oder Biomolekülen (Nukleinsäuren - DNS, RNS - und verschiedenste Proteingruppen) verstanden wer
den, die durch Zerlegung von Organismen in funk
tionale oder stoffliche Bestandteile gewonnen (isoliert, gereinigt etc.) werden können. Zum an
deren bedarf sie der Etablierung einer technischen Bearbeitungsstruktur, in die Isoliertes und Extra
hiertes eingeführt und mit gegenüber seinem pri
mären Zusammenhang autonomen Verfahren pro
zessiert werden kann.
Diese Voraussetzungen sind in den untersuchten Laboratorien gegeben. In den beobachteten Ex
perimenten besteht eine weitgehende Indifferenz der verwendeten allgemeinen gentechnischen Ver
fahren gegenüber der Herkunft des Materials aus verschiedenen Organismen. Für die Verfahren sind Fragen der Verfügbarkeit, der Leichtigkeit des Gewinnens, der Stabilität und der Reinheit der Materialien vorrangig vor den Details ihrer Herkunft.15
Sichtbar wird die Autonomie auch in der techni
schen Identität der Laborordnungen in unter
schiedlichen Laboratorien, die unabhängig von den konkreten Organismen, die die ’Lieferanten4 des Materials sind, verfügbar sind. Die Erzeugung und Erhaltung einer massiven Infrastruktur durch ein Netz von Laboratorien ist das Resultat der bio-
15 Damit soll selbstverständlich nicht behauptet wer
den, daß die Wahl des Materials für die konkreten Wissensprozesse irrelevant wäre.
technischen Innovationen der letzten fünfzig, ins
besondere aber der letzten zwanzig Jahre.16 Mit ihnen hat ein Laboratorisierungsschub biologi
scher Forschung auf breiter Front stattgefunden.
Wir haben es mit einer doppelten Autonomie der Verfahren zu tun: sie sind zum einen als technische Verfahren in Prozeduren und Apparaturen stabili
siert; und sie sind zum anderen in ihrer Verwen
dung in vielfältiger Hinsicht unabhängig von den Unterscheidungen zwischen den Objektklassen der Biologie. Als die in molekularbiologischen La
boren arrangierten analytischen Mitteln bilden sie einen stabilen apparativ-technischen Rahmen, in dem sich nun weitere Strukturierungen bestimmen lassen.17
3.2 Modellsysteme, Biotope und Laboratope In den Laboren stoßen wir u.a. auf routinisierte Sprechweisen, mit denen unterschiedliche Phäno
mene der Forschungspraxis als ’Modellsystem4 charakterisiert werden. „Mein Modellsystem ist F944 oder „Dazu verwende ich das Reticolozyten- System“ , oder: „In unserem Modellsystem ist das sehr schwierig darzustellen44 sind Beispiele solcher Sprechweisen, die sich in vielfältiger Weise im eth
nographischen Datenmaterial zeigen.
Modellsysteme erlauben offenbar theoretische und praktische Operationen als deren Gemein
samkeit angesehen werden kann, daß sie an denje
nigen Objekten, für die sie ein Modell sind, entwe
der nicht möglich sind oder zu aufwendig wären.
So wird die Maus als das Modellsystem für die U n
tersuchung der Säugetierentwicklung und damit letztlich für die Entwicklung des Menschen, die in vitro Differenzierung von Zellen, die als Zellinien in Laboren kultiviert werden als das Modellsystem für die Mausentwicklung, eine isolierte und gerei
nigte DNS in einem Transkriptionssystem als das Modell für Differenzierungsprozesse in Körperzel
len betrachtet.
Ist das Modellsystem ’die Maus4, so wird diese Maus als ein ’natürlicher4 Ersatz für die - aus ver
schiedensten Gründen nicht zu verwendenden - Menschen und als Prototyp für Säugetiere verstan-
16 Einen kurzen Überblick über die zentralen Innova
tionen in der Molekularbiologie mit den Hinweisen auf die zugehörigen Fachpublikationen seit 1938 gibt Witkowski (1988).
17 Zu den wissenssoziologischen Eigenheiten dieses Rahmens vgl. Amann (1990).
28 Zeitschrift für Soziologie, Jg. 23, Heft 1, Februar 1994, S. 22-40
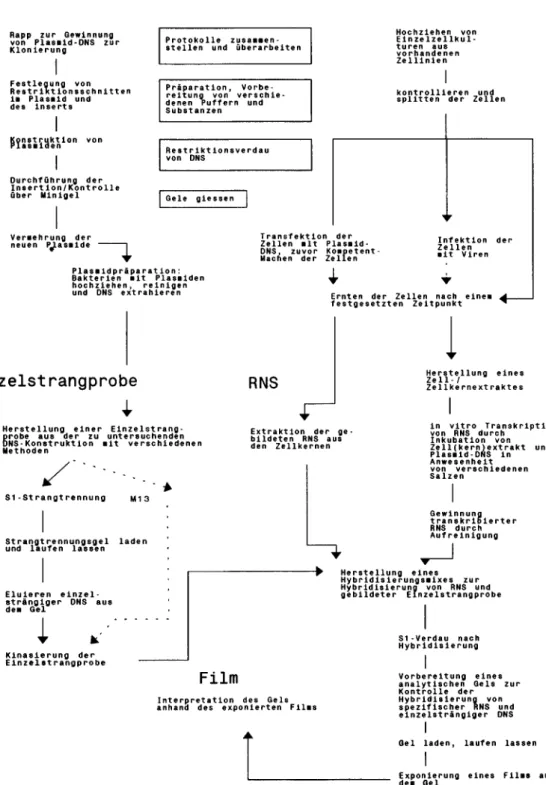
Ausgangsmaterial Zellen
Rapp zur G e wi nn un g von Pl as ai d- DN S zur K l o n i e r u n g
P r o t ok ol le z u s a a a e n - stellen und üb er ar be it en
H o ch zi eh en von E i n z e l z e l l k u l - ture n aus v o r h a n d e n e n Z e l l i n i e n Fe st l e g u n g von
R e s t r i k t i o n s s c h n i t t e n ia Plasaid und des insert s
D u r c h f ü h r u n g der I n s e r t i o n / K o n t r o l l e über Mi ni g e l
Ve rie h
neuen rung der ''lasaide
P r ä p a r a t i o n , V o r b e - reituno1 von verschie- denen 1Puffer n und S u b s t a n z e n
k o n t r o l l i e r e n und splitten der Zellen
R e s t r i k t i o n s v e r d a u von DNS
Gele gi essen
T r a n s f e k t i o n der Ze llen alt Plasald- D N S , zuvor Koapetent- Machen der Zellen
In fe kt io n der Z e l l e n ait Viren P l a s a i d p r ä p a r a t i o n :
Ba kt er ie n ait Plas ai de n ho chz ieh en , re in ig en und DNS ex tr ah ie re n
4 V
Ernten der Zellen nach einea f e s t g e s e t z t e n Ze it pu nk t
Einzelstrangprobe RNS
H e r s t e l l u n g eine s Z e l l - /Z e l l k e r n e x t r a k t e s
H e r s t e l l u n g e i n e r E i n z e l s t r a n g probe aus der zu u n t e rs uc he nd en D N S - K o n s t r u k t i o n ait v e r s c h i e d e n e n M e t h o d e n
S 1 - S t r a n g t r e n n u n g M 1 3 *
Ex tr ak ti on der g e bildeten RNS aus den Ze ll ke rn en
in vitro T r a n sk ri pt io n von RNS durch
In ku ba ti on von Z e l l (k e r n )ex tr a k t und P l a s a i d - DNS in A n w e s e n h e i t von v e r s c h i e d e n e n S a l z e n
S t r a n g t r e n n u n g s g e l la de n und la uf en la sse n
E l u i e r e n ei nz el - s t r ä n g i g e r DN S aus den Gel
K i n a s i e r u n g de r
i .
E i n z e l s t r a n g p r o b e
G e w i n n u n g t r a n s k r i b i e r t e r RNS durch A u f r e i n i g u n g
4 T__I
H e r s t e l l u n g eines H y b r i d l s i e r u n g 8 a i x e s zur H y b r id is ie ru ng von RNS und g e b i l d e t e r E i n z e l s t r a n g p r o b e
S1 -V e r d a u nach H y b r i d i s i e r u n g
Film
In te rp re ta ti on des Gels anhand des ex po ni er te n Files
V o r b e r e i t u n g eines an al yt i s c h e n Ge ls zur Ko nt r o l l e der H y b r i d i s i e r u n g von sp ez if i s c h e r RN S und e i n z e l s t r ä n g i g e r DNS
I
Ge l laden, laufen lassen
Ex po ni er un g eines Filas auf dea Gel
Abb. 1 Prozessierung von biologischem Material (schematisch).
Klaus Amann: Menschen, Mäuse und Fliegen 29 den.18 Zu zeigen ist hier, daß dieses Repräsenta
tionsverhältnis zwischen verschiedenen biologi
schen Objekten, der Modellcharakter, lediglich ei
nen Darstellungsaspekt der jeweiligen Teilbereiche der Molekularbiologie zeigt. Handelte es sich bei diesen Modellsysteme lediglich um eine - mögli
cherweise vorübergehende - technische oder sozi
ale Schwierigkeiten überbrückende Forschungs
strategie, mit der ersatzweise andere Organismen statt der ’eigentlich’ interessierenden in ihre mate
rialen Komponenten zerlegt werden, so ergäbe sich daraus keine systematisch neue Perspektive auf die Gegenstände der Biologie.
Neue Struktureinheiten werden Modellsysteme je
doch als instrumentell-technische Rekonstruktio
nen oder Rekonfigurationen der biologischen Ge
genstände. Ich betrachte sie als komplexe Einhei
ten von experimentellen Modellierungsprozessen im Labor und nicht als natürliche Repräsenta
tionssysteme. Das biologische Labor gewinnt mit ihnen jenseits seiner Eigenschaft als ein techni
sches Arrangement zur Dekonstruktion von Lebe
wesen, das zur Produktion eines Wissens um deren Eigenschaften und Funktionsweisen genutzt wer
den kann, die Bedeutung einer zweiten Natur. Es tritt als Bezugsgröße der Wissensproduktion an die Stelle einer ’natürlichen N atur4, in der wir noch mit vertrauten oder unvertrauten Organismen ei
ne Lebenswelt bewohnen.
In dieser Lebenswelt, in der wir uns etwa als Teil der Biotope eines Wohnhauses begreifen können, das neben uns die Mäuse im Keller, die Fliegen an der Lampe, die Bakterien im Darm und die Viren im Krankenbett umfassen kann - befinden sich die genannten Organismen in einem Zustand, den man gegenüber der zweiten Natur des Labors als primären Produktionszusammenhang biologi
schen Materials beschreiben könnte.19 Nicht nur die Organismen, sondern auch das Wissen und un
sere Erfahrungen aus der Mitwohnerschaft mit den genannten Lebewesen wird bei der Gewin
nung von biologischem Material nahezu irrelevant und verschwindet mit der Lebendigkeit der getöte
ten Organismen aus der Wissenschaft vom Leben
digen.
18 Dafür gibt es eine Reihe ausformulierter theoreti
scher wie forschungstechnischer Gründe: z.B. die evolutionäre Nähe der Maus zum Menschen einer
seits, die leichte Handhabbarkeit der Maus gegen
über anderen Säugern andererseits.
19 Dieses Beispiel soll deutlich machen, daß mit primär weder ’ursprüngliche* noch ’wilde4 Natur oder ähnli
ches assoznert werden sollte.
Die zweite Natur im Labor bildet die lokal stabili
sierte Umwelt für den epistemologisch bedeutsa
men Übergang von der Maus als lebensweltlichem Organismus zu einem Modellsystem mit dem Na
men Maus. In den letzten Zuckungen eines getöte
ten Tieres liegen die letzten sichtbaren Ähnlichkei
ten biologischen Materials mit dem lebensweltlich erkennbaren Tier. Die entscheidenden Möglich
keiten jeder analytischen labor-wissenschaftlichen Bearbeitung beginnen mit dem Ende des Lebens:
Lebendigkeit ist insbesondere für die epistemi- schen Objekte der modernen Biologie eine vor
übergehende Erscheinung.
Aber nicht erst der Tod und die Zerlegung markie
ren den Bruch, durch den sich der Objektcharak
ter von Labortieren und anderen Tieren unter
scheiden. Auch wenn die in lebensweltlichen oder
’natürlichen* Konfigurationen wie dem eben be
schriebenen Biotop des Hauses vorkommenden,
’Hausmäuse* genannten Mitbewohner dem unbe
darften Besucher eines Labors noch als Verwandte der Labormäuse erscheinen mögen, so sind sie dies für uns in keinem wissenssoziologisch vernünf
tigen Sinn mehr.
Die Unterstellung einer Verwandtschaftsbezie
hung zwischen der Maus im Keller und der Maus im Laborstall verkennt gerade die wissenschaftli
chen Rekonfigurationsleistungen der Laborbiolo
gie, die beide voneinander trennt und sie wechsel
seitig als fundamental unterschiedene Objekte konstituiert. Die Identifizierung beider Objektty
pen miteinander ist bestenfalls das Resultat einer theoretischen Operation, die praktische Nichtun
terscheidbarkeit unter Umständen das Resultat ei
ner Unaufmerksamkeit der Tierpfleger oder Wis
senschaftler, die einen Stall offengelassen haben.
Plakativ gesagt: Natur im ursprünglichen Sinne des Wortes hat als Gegenstand keinen Platz (mehr) innerhalb der Grenzen des Labors.20 Modellsyste
me haben als ihr strukturelles Resultat die Auto- nomisierung von Lebewesen aus ihren bisherigen Existenzbedingungen, deren Rekonfiguration als epistemische Dinge, und ihre dauerhafte Erhal
tung als epistemische Objekte innerhalb von loka
len Labor-Infrastrukturen, die eine zweiten Natur etablieren.
Mein Bild des molekularbiologischen Labors zeigt nun zweierlei: auf der einen Seite das komplexe 20 Was nicht heißt, daß man in Laboren keine Haus
mäuse finden könnte. Ich habe selbst meine erste Maus im Labor, eine Hausmaus, unter einem Schreibtisch am Butterbrot eines Wissenschaftlers nagen sehen.
30 Zeitschrift für Soziologie, Jg. 23, Heft 1, Februar 1994, S. 22-40 Arrangement einer technischen Infrastruktur zur
Gewinnung und Analyse biologischen Materials.
Auf der anderen Seite eine Autonomisierung der Bezugsobjekte des Wissensprozesses, die zugleich die Konstitution lokal prozessierter neuer biologi
scher Objekte darstellt.
Um dieses lokale Prozessieren zu ermöglichen, werden Räume innerhalb von Laboren etabliert, die ich im Unterschied zu Biotopen als Laboratope bezeichnen will.21 Laboratope sind Umwelten für die neuen Objekte, die Technofakte der Biologie, die innerhalb von Laboratorien erzeugt werden.22 Es geht nun nicht mehr nur darum, daß einzelne . Lebewesen technisch verändert oder Teile von ih
nen modifiziert und isoliert behandelt werden können. Diese neuen Objekte sind vielmehr Ele
mente in wissenschaftlich-technisch durchstruktu
rierten Umwelten. Sie sind beispielsweise nur dann und nur so lange ’lebendig4 wie innerhalb von Laboratorien entsprechende technische Arrange
ments aufrechterhalten werden.
Man könnte nochmals einwenden, daß die Identi
tät der Lebewesen im Labor nach wie vor gewahrt bleibt: in den Tierställen sind Lebewesen versam
melt, deren Vorfahren irgendwann ’wild4 gelebt hatten. Sie werden gezüchtet und ’verbraucht4.
Züchtung ist - zumindest für unsere Gesellschafts
form - eine auch lebensweltlich etablierte Praxis, die hier lediglich für andere Zwecke und an ande
ren Lebewesen optimiert erscheint.23 * Diese Beob
21 Dabei will ich zugunsten der begrifflichen Gegen
überstellung die Bildung eines Sprachhybrids aus griechischen und lateinischen Stämmen hinnehmen.
22 Ich betrachte hier lediglich diejenigen Entwicklun
gen innerhalb der Molekularbiologie, die gegenüber einem ’Rücktransport4 ihrer neuen Objekte in Bioto
pe gegenwärtig indifferent sind. Allerdings vermute ich, daß der zentrale Punkt in der laufenden öffentli
chen Debatte über die ’Freisetzung gentechnisch ver
änderter Organismen4 mit der von mir unternomme
nen Unterscheidung zwischen ’Organismen4 und bio
logischen ’Technofakten4 genauer bestimmt werden könnte.
23 Die Tierzucht als Grundlage menschlicher Ernäh
rung hat in Form der modernen Tierstallproduktion einige Parallelen zum Laboratop der Wissensproduk
tion. Auch hier werden natürliche Organismen über gezielte Mechanismen der Zuchtwahl und der Orga
nisation des Tierstalles (Ernährung, Beleuchtung etc.) zu Produktionsstätten von Lebensmitteln und erlangen eine zweite Natur als Produktionsmittel.
Siehe auch Latours Arbeit über Pasteur (1988) in der die Verwissenschaftlichung der Tierhaltung durch Laboratorisierung analysiert wird.
achtung scheint auf einen unveränderten Kern von natürlichem und Labortier hinzuweisen: fortbeste
hende Lebendigkeit. Die weitere Beobachtung er
gibt ein anderes Bild, das den Bezug zwischen bei
den Objektformen schrittweise auflöst. Am Ende des Auflösungsprozesses steht als ein Resultat die Rekonfiguration von Lebewesen als epistemische Objekte, als Produkte und Träger wissenschaftli
chen Wissens.
Um von Organismen zu diesen neuen Objekten zu kommen und in lokalen Wissensprozessen über die dafür notwendigen stabile Infrastrukturbedingun
gen zu kommen, bedurfte es einer Reihe von R e
konfigurationsschritten im Umgang mit ’natürli
chen4 Organismen. Sind für die jeweiligen Lebe
wesen vielfältig und unterschiedlich weit gediehen und könnten sowohl disziplingeschichtlich als auch einzelfallorientiert rekonstruiert werden.
Ich will die wichtigsten für das hier weiter zu ver
folgende Beispiel ’Maus4 zumindest nennen. Es sind infrastrukturelle Rahmenbedingungen, die heute als gegeben angesehen werden können:
• die Systematisierung der Züchtung durch pro
fessionelle und kommerzielle Labors
• die Erzeugung von ’reinen4 Rassen durch Züch
tungsverfahren
• die Standardisierung von Haltungsbedingungen
• die Dokumentation von Forschungsergebnissen an und mit Varianten
• die Erhaltung und ’Pflege4 der Varianten
• die rigorose Kontrolle des Zugangs von ’neuem 4 Material in die forschungsinterne Prozessierung.
Die Maus ist wohl das mit Abstand am besten un
tersuchte Säugetier. Umfangreiche Literatur fin
det sich zur Entwicklung oder zur Genetik der Maus; ebenso zur Haltung, zur Manipulation, zur Züchtung. Ohne Maus keine Reproduktionsmedi
zin, keine Tests medizinisch-pharmazeutischen Wissens und Könnens. A ber eine Integration ’wil
der4 Mäuse oder Hausmäuse, ihr Transport in lo
kale Laboratope ist meines Wissens nirgendwo Praxis in Laboren, die wie die Molekularbiologie an Mäusen Forschung betreiben. Was wir dagegen finden sind Mäusefabrikationsstätten wie etwa das US-amerikanische Jackson Labor oder internatio
nal operierende Firmen, die über Kataloge Mäuse als käufliche Ware anbieten, entweder als Lebend- M aterial oder als DNS-Substanz. In deren Katalo
gen sind verschiedenartigste Produkte im Angebot und die jeweiligen Produkteigenschaften spezifi
ziert. Selbst über Waren, die nicht (mehr) herge
stellt werden, finden wir Informationen etwa: ’aus
gestorben4. Diese Produktionsstätten sindTeü der
Klaus Amann: Menschen, Mäuse und Fliegen 31 Infrastruktur (Ökologie und Ökonomie) von
Laboratopen.24
Neben diesen Produktionsstätten gehört als ein weiteres Element die Tätigkeit von Klassifikations
institutionen zur äußeren Infrastruktur der mole
kularbiologischen ’Mausforschung*. Diese sorgen für die Einhaltung und Weiterentwicklung der Be
nennung von Bestandteilen des genetischen M ate
rials, das weltweit in Laboren analysiert, auf der genomischen Karte der Maus lokalisiert und dann allgemeinverbindlich eingetragen wird.
Das Labortier ’Maus*, dem wir heute in For
schungslaboratorien begegnen, ist demnach schon immer ein wissenschaftliches Produkt, das einen Namen bzw. eine Stamm-Nummer und eine Viel
zahl forschungsrelevanter Eigenschaften besitzt, die ihm abgerungen und/oder zugefügt wurden.
Das gilt auch für die ’Wildtyp-Maus*, die als Kon
trollorganismus Verwendung findet und keinesfalls irgendeine von Wissenschaftlern im Institutskeller gefangene Maus ist, sondern eine wohldefinierte Rasse bei deren Selektion u.a. auf ihre Pflege
leichtigkeit unter Laborbedingungen geachtet wurde.
3.3 Die lokale Etablierung von Laboratopen und ihre Einbettung in eine analytische
Infrastruktur
Die bisherigen Ausführungen bezogen sich vorran
gig auf infrastrukturellen Voraussetzungen für die jeweils lokalen Möglichkeiten der Teilhabe an der Erforschung von Objekten einer zweiten Natur.
Anders gesagt: in der Schaffung und Erhaltung dieser Infrastruktur stecken die Bedingungen, daß nicht mit beliebigen zweiten, ideosynkratischen
’Naturen* inkommensurable Erkenntnisse gene
riert werden. Um auf der Ebene der Wissenspro
duktion mit diesen neuen Objekten zu Resultaten zu kommen, für die Geltung beansprucht werden kann, muß auf lokaler Ebene ein funktionsfähiges Arrangement von Laboratopen und analytischer Infrastruktur erzeugt werden.
Als Gründungspaar einer Mauszucht im eigenen Haus (Forschungsinstitut) bedürfen die dafür er
worbenen Exemplare einer Reihe von Behandlun
gen, um sie zu den Ureinwohnern eines Labora- tops innerhalb eines konkreten Labors zu machen:
den Abschluß gegen eine verschmutzende Umwelt einschließlich aller Personen im Labor, eine lokale 24 Bislang meines Wissen nicht Gegenstand soziologi
scher Untersuchung.
Tierstallinfrastruktur mit Pflege, Zucht, Buchhal
tung, Beauftragtem für den Tierschutz etc.
Die auf einen oder wenige Stämme mit ’homoge
nisiertem genetischem background*25 konzentrier
te Tierhaltung schafft die lokale Voraussetzung für die Kontrolle wichtiger Randbedingungen des weiteren experimentellen Geschehens. Jedes wei
tere Ergebnis, das mit dem darauf errichteten lo
kal spezifizierten System gewonnen werden kann, ist auf den Nachweis angewiesen, daß die erzeug
ten und beschriebenen Effekte keine Effekte einer von bestimmten Normierungen abweichenden ge
netischen, hygienischen oder gar epidemiologi
schen Ursache - der ’Durchseuchung eines Tier
stalls* mit Krankheitserregern - sind. Die ’Kon- trollmaus* (häufig die Wildtyp-Maus) muß ebenso wie die untersuchte Maus ein immer wieder mit ei
nem Standardtyp hinreichend identisches Exem
plar darstellen.
Die Produkte (’Bewohner*) der Laboratope ste
hen in einer systematischen Wechselwirkung mit den analytischen Bearbeitungsverfahren biologi
schen Materials. Sie werden dabei als Material be
nutzt und verbraucht.26 Sie werden nicht nur in ei
nem theoretischen Sinne in ihre Bestandteile auf
gelöst, sondern sind das Ausgangsmaterial, das nun immer wieder neu gewonnen werden kann und muß, um gentechnische Untersuchungen durchzuführen. Dabei kommt es zu einer weiteren Rekonfiguration im Prozeß ihrer Analyse, die die ursprünglichen Bewohner nun zu Repräsentanten experimenteller Effekte und Resultate macht.
In den beobachteten Laboren geht es in weiten Bereichen um zwei Klassen von Materialien, die aus Mäusen (aber auch Zellen und Bakterien) prä
pariert werden: sogenannte Schnittpräparate (nur von Mäusen) und DNS- bzw. RNS-Fraktionen.
Schnittpräparate sind das Produkt von getöteten Mäusen oder Mausembryonen, die mit Schneide
apparaturen in hauchdünne Scheiben geschnitten
25 Mit diesem Teilnehmerbegriff ist u.a. die Praxis der Etablierung von sogenannten Inzuchtstämmen ge
meint, deren genetische Varianz durch Züchtung mi
nimiert wurde.
26 Die ethische Problematik dieses Umgangs mit Lebe
wesen erscheint mir aus meinen Beobachtungen durchweg mit einem überaus hohen Maß an Verant
wortung und Bewußtsein für das einzelne Tier ver
knüpft: es gibt systematische Kontrollen und Überle
gungen, das Ausmaß der Verwendung von Tieren so gering wie möglich zu halten. Dies wird diejenigen allerdings kaum beruhigen, die Tierversuche per se für verwerflich halten.
32 Zeitschrift für Soziologie, Jg. 23, Heft 1, Februar 1994, S. 22-40
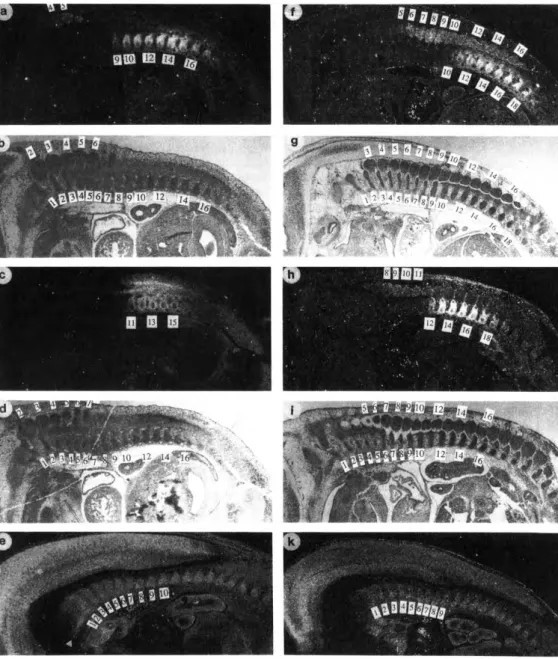
Figure 6. Hox Gene Expression in RA-Exposed Embryos
Day 12.5 embryos exposed to RA on day 7, 4 hr (a-e) or unexposed (f-k) were analyzed as described in the text, (b), (d), (g), and (i) are bright-field exposures of (a), (c), (0, and (h), respectively. The probes were specific for the following genes: Hox-1.1, (a) and (f); Hox-3.1, (c) and (h); and Hox-1.5, (e) and (k). Spinal ganglia and prevertebrae are numbered; in the dark-field exposures the first number indicates the anterior boundary of expression.
The arrowhead in (e) points out the RA-induced Hox-1.5 expression domain ventral to the basioccipital bone anlage. Note the anterior shifts of expression domains induced by RA exposure as discussed in the text.
Abb. 2 E m b r y o s c h n i t t e - B i l d e r a u s e i n e r m o l e k u l a r b i o l o g i s c h e n P u b l i k a t i o n ( a u s : C e l l , V o l . 6 7 : 9 5 , O c t . 1 9 9 1 ) .
34 Zeitschrift für Soziologie, Jg. 23, Heft 1, Februar 1994, S. 22-40 Strukturen zur Stabilisierung einer zweiten Natur,
die Unterhaltung von lokalen Laboratopen als Voraussetzung zur Teilhabe an den dazugehörigen Wissensprozessen.
Daraus entstehen in den Laboren Einheiten, die im wissenssoziologischen Sinne Modellierungssy
steme sind. In ihnen werden z. B. Mäuse als episte- mische Objekte modelliert. Es sind Systeme, in denen aus einzelnen Mäusen Träger theoretischer Eigenschaften werden. Sie sind nicht mehr nur pas
sive ’natürliche’ Objekte, die in experimentellen Arrangements etwas von ihrem inneren Zusam
menhalt preisgeben. Sie bringen als Resultat ihrer Modellierung vielmehr ein theoretisches Konzept zur körperlichen Erscheinung und liefern sich selbst als Material. Sie werden in ihrer zu (visuel
len) Repräsentationen aufbereiteten Form zu em
pirischem Beweismaterial.
D am it Mäuse (oder andere H ere) diese Leistung vollbringen können, müssen sie in den skizzierten Hinsichten transformiert und in ein technisches System eingebettet werden. Dazu gehört, daß sie in eine praktische Relation zu den Bewohnern an
derer Laboratope (meist innerhalb desselben La
bors) gebracht werden. Mäuse sind in den betrach
teten Forschungskontexten ein Objekttypus, der in einem Laboratop eine neue Identität erhält und gewinnt. Daneben gibt es beispielsweise als soge
nannte Zellinien etablierte Körperzellen unter
schiedlicher Herkunft und insbesondere Bakte
rien, die Forschungs- und Produktionsaufgaben übernehm en müssen. Mäuse werden zu einer Ka
tegorie von Bezugsobjekten der Forschung. Inner
halb von Modellierungssystemen liefern sie M ate
rialien zu ihrer eigenen Erforschung. Andere La
boratope im Kontext der untersuchten Labore be
herbergen zelluläre oder bakterielle Systeme. D ie
se bakteriellen Systeme etwa können so ins Labor integriert sein, daß sie selbst nicht vorrangig der
’eigentliche4 Gegenstand der Forschung sind.29 Sie sind dann die mit gentechnischen Mitteln systema
tisch veränderten lebendigen Fabriken oder Pro
duktionssysteme, mit denen Material, beispiels
weise Nukleinsäuren (genomische Sequenzen) aus der Maus prozessiert werden können. Sie könnten im Prinzip ebenso Sequenzen aus menschlichen oder anderen tierischen Organismen verarbei-
29 Was nicht bedeutet, das ’dieselben4 Bakterien in an
deren Kontexten nicht doch der Kern eines Modell
system sein könnten: etwa für den Test der Funktion einer Nukleinsäuresequenz, die selbst aus einem an
deren, z.B. tierischen Organismus stammt.
ten.30 Die für diese Systeme notwendigen Labora
tope werden mithilfe von Gefrierschränken, Warmräumen, Nährlösungen und Schüttelkolben etabliert, in denen die jeweiligen Exemplare mas
senhaft bevorratet, angezüchtet, vervielfältigt und zum A rbeiten gebracht werden: zur Vervielfälti
gung und Transformation von genetischem M ate
rial.31 Ihre Bewohner fristen ein kontrolliertes D a
sein innerhalb vollständig artifizieller Laboratope.
Sie sind eingebunden in eine Forschungsökologie und -Ökonomie, die neue Lebensfunktionen von den Bewohnern verlangt: z.B . die Produktion
’fremder4 Substanzen, eine entgrenzte Lebensfä
higkeit, die Konzentration auf eine Lebensfunk
tion, die Suspendierung von evolutionären Mecha
nismen bzw. die Instrumentalisierung isolierter Mechanismen wie etwa Selektionsdruck.
Diese Laboratope sind hochempfindliche, künstli
che Produktionssysteme. Hilfsmittel zu ihrem Er
halt sind biologische und biochemische Substan
zen, die als biologische Werkzeuge zur Aufrechter
haltung des Laboratops, aber auch zur systemati
schen Dekonstruktion seiner Bewohner genutzt werden können.
Ihre Unterscheidbarkeit als Exemplare mit redu
zierten, arbeitsteilig organisierten und exakt zuge
schnittenen Eigenschaften macht sie zu keinen neuen Arten im Sinne der klassischen Systematik.
Die neuen taxonomischen Ordnungsmuster, die für diese biologischen Technofakte etabliert wur
den, sind vielmehr technischen Spezifikationen vergleichbar wie man sie beispielsweise für Ele
mente elektronischer Schaltungen in der Elektro
technik kennt. Ihre Individualität und Identität er
gibt sich einerseits in rudimentärer Weise aus dem Ursprungsstamm eines Bakteriums, andererseits aber aus einer forschungstechnischen Bio
graphie.32 * Sie haben eine lokale Nummer, einen Konstrukteur, eine Referenz und Spezifikationen ihrer Haltungsbedingungen. (Vgl. Abbüdung 4) 30 Im Prinzip heißt, daß es natürlich sehr unterschied
lich konstruierte Fabrikationssysteme sind, die je
weils biologisches Material produzieren. Bekannte
stes Produkt solcher Systeme ist das biotechnisch hergestellte Human-Insulin.
31 Neuere Entwicklungen, deren Erfinder 1993 mit dem Nobelpreis honoriert wurden, substituieren teil
weise diese lebendigen Fabriken durch biochemische Prozeßautomaten: PCR-Maschinen.
32 Wobei auch hier wiederum güt, daß es sich um nur wenige unterschiedliche Stämme handelt, die durch Generationen von Laboratorien domestiziert, sele- giert und buchstäblich auf elementare Teüe ’zurück
gestutzt4 wurden.
Klaus Amann: Menschen, Mäuse und Fliegen 35
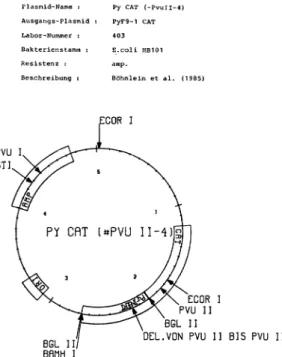
rlasmid-Name : Py CAT (-PvuII-4) Ausgangs-Plasmid : PyF9-1 CAT
Labor-Nummer : 403
Bakterienstamm : E.coli HB101
Resistenz : amp.
Beschreibung : Böhnlein et al. (1985)
Abb. 4 Plasmidkarte - lokale Datenbankeinträge für ein gentechnisches Instrument.
In dieser Form werden sie als ’Werkzeuge4 charak
terisiert und in Datenbanken registriert, viele auch als kommerzielle Systeme in Katalogen beschrie
ben und zum Kauf angeboten. In ihrer Existenz sind sie vollständig abhängig von der Sorge der La
boratorien und ihrer Produzenten. Sie sind nicht rückführbar in lebensweltliche Biotope.33
An diesem Beispiel wird deutlich, daß der Produk
tionsaspekt und der Modellierungsaspekt von Technofakten voneinander unterscheidbar werden.
Die Voraussetzung für die kontinuierliche Produk
tion biologischen Materials mit (bio-)technischen Verfahren liegt im Übergang von einem ursprüng
lich, d. h. im Konstitutionsprozeß epistemisch defi
nierten und etablierten Ding zu materiellen Objek
ten, zu technischen Apparaturen. Die Besonder
heit dieser Materialisierungen ist, daß sie das R e
sultat eines Transformationsprozesses sind, in dem sie über eine Phase der Abtrennung und Aufspal
tung ’natürlicher4 Einheiten zum Kern von Model
lierungssystemen und damit zu Gegenständen von Wissensprozessen werden, aus denen schließlich re- konfigurierte technische Einheiten (Technofakte) 33 Hoffentlich nicht rückführbar - wie u.a. auch die
Kritiker der Gentechnik einfordem.
entstehen, die in eine wissensbasierte Produktions
logik eingebettet sind und als materiale Wissenspro
dukte Realität gewinnen.
Mäuse erscheinen gegenüber Bakterien als un
gleich komplexere Organismen, deren Transfor
mation in ein nahezu vollständig kontrollierbares technisches Ding gegenwärtig unwahrscheinlich erscheint.34 Die Erzeugung von Produktionssyste
men, die neben dem Laboratop der Maus noch weitere Räume innerhalb von Laboren strukturie
ren, macht es jedoch möglich, über eine temporä
re, vollständige Entleiblichung biologischen M ate
rials aus der Maus und durch seine Prozessierung in artifiziellen Laboratopen wie denen von bakte
riellen Systemen eine neue Qualität biologischen Wissens zu etablieren. Diese entsteht u.a. da
durch, daß technische Möglichkeiten entwickelt wurden, mit denen über die Grenzen von Arten und Klassen hinweg nicht nur Merkmale und Rea
lisierungen von Lebensfunktionen verglichen wer
den können, sondern auch ’natürliche4 Merkmals
differenzen von Lebewesen in experimentell-tech
nische Instrumente zur Wissensproduktion trans
formiert werden können. Menschen, Mäuse und Fliegen können nicht mehr nur verglichen, sie können zur wechselseitigen materialen Ressource ihrer Erforschung gemacht werden.
So haben beispielsweise zu Beginn der achtziger Jahre Drosophila-Genetiker - nun ausgestattet mit den technischen Möglichkeiten der Molekularbio
logie und nicht allein der klassischen Genetik - die (Teilstücke der) DNS eines Genes identifizieren und isolieren können, dessen Defekt eine Desor
ganisation der Fliege erzeugt.35 Dieses Gen(stück) konnte dann als in ein Bakterium integriertes DNS-Stück durch andere Forschungslaboratorien bezogen werden. Gentechnische Verfahren er
möglichten es zu diesem Zeitpunkt, beliebige DNS-Stücke als Detektoren oder Sonden für die Suche nach ähnlichen Sequenzen in verschieden
sten Organismen zu verwenden. Vorausgesetzt, man gehörte zu denjenigen, die diese Sonde beka
men und die zugleich über ein Laboratop verfüg
ten, in dem die gerade beschriebenen Verfahren einer Extraktion von genetischem Material aus ei
nem bestimmten Organismus bereits funktionier- 34 Wenngleich auch ein Biologe mir gegenüber genau solche (für ihn überaus bedenkliche) Visionen als Perspektive dieser Molekularbiologie formuliert hat.
35 Vgl. dazu eine populärwissenschaftliche Darstellung des Entdeckers W. Gehring in Bild der Wissenschaft (1984) oder den Fachartikel von McGinnis et al.
(1984a).
36 Zeitschrift für Soziologie, Jg. 23, Heft 1, Februar 1994, S. 22-40 ten, konnte nun die Suche beginnen. Im April 1985
erschien, als ’article4 in Nature veröffentlicht, eine Publikation, die den Erfolg dieser Strategie de
m onstrierte.36
Die dazugehörige experimentelle Arbeit vollzieht einen wichtigen und großen Schritt weg von einer Beschränkung der molekularbiologischen For
schung in den Grenzen ’natürlicher’ A rten, hin zu einer experimentellen wie konzeptuellen Transfor
mation von unterschiedlichen Tierarten zu Model
lierungssystemen. Aus einer verglichen etwa mit der Erfindung von Nukleinsäure-Sequenzierungs
techniken unscheinbaren Erweiterung technischer Möglichkeiten folgen aber auch disziplinäre, die soziale Organisation der Wissensprozesse verän
dernde Konsequenzen.
Es verändert sich also nicht nur der Gegenstand der jeweiligen Spezialisten von einem ’externen4 oder ’natürlichen4 Objekt zu einem Träger episte- mischer Eigenschaften, auch subdisziplinäre Grenzziehungen entlang den ’natürlichen4 Einhei
ten wie Maus, Fliege oder Hefepilz werden obso
let. Die A rbeit mit und an einem bestimmten Orga
nismus ist nun in einer neuen Weise zugleich von praktischer Relevanz und Beliebigkeit. Relevant wird sie dann, wenn dieser Organismus den Kern eines lokal etablierten Modellierungssystems bil
det. Dieses System kann dann zum A rbeiten ge
bracht werden und dazu dienen, mit vielfältigen Importen aus sehr unterschiedlichen Zusammen
hängen neue Erkenntnisse zu produzieren.
Entscheidend wird es für die Forschung, über ein Arrangem ent von Laboratopen zu verfügen, mit dessen Hilfe biologisches M aterial erzeugt, pro
zessiert und die Resultate in einem lokal handhab
baren Modellsystem integriert werden können.37 * * * * * * Daraus ergeben sich veränderte Kommunikations
und Interaktionsbeziehungen, die nun nicht mehr vorrangig entlang der traditionellen Differenzie
rungen der Biologie verlaufen, sondern an geneti
schen M ustern, ihren Ähnlichkeiten und Differen
zen entlang entstehen. Die technischen Eigenlogi
36 Colberg-Poley (1985), kurz zuvor McGinnis (1984b).
’Article4 in NATURE bedeutet in der community die höchste Priorität einer Veröffentlichung und steht je
weils für einen zentralen Beitrag zur Weiterentwick
lung der Disziplin.
37 Bei genauerer Betrachtung unterschiedlicher Mo
dellsysteme wird deutlich, daß es für deren Wahl komplexer Entscheidungsprozesse bedarf und zum Zeitpunkt einer Entscheidung für ein bestimmtes Sy
stem keine genaue Perspektive seiner Leistungsfä
higkeit angegeben werden kann. Dies kann hier nicht weiter verfolgt werden.
ken der jeweiligen Systeme sind an der Bearbei
tung von bestimmten Materialien orientiert, deren Eigenschaften weitgehend unabhängig von kon
kreten Organismen erscheinen. Die Einführung der Gentechniken hat zu einer großen Vereinheit
lichung der technischen Infrastrukturen geführt:
„Du kannst überall auf der Welt hingehen. Die mo
lekularbiologischen Labore sehen alle gleich aus44, so ein Laborleiter im Gespräch.
Die Unterschiede, und damit auch verschiedene Zugangsmöglichkeiten und Teilhabe von Forscher
gruppen an der Forschungsfront ergeben sich nun nicht allein aus den materialen Ressourcen der La
bore oder dem Wissen um die Besonderheiten ei
nes für die Fachwelt interessanten Organismus oder bestimmter Funktionskomplexe. M itent
scheidend wird, daß für einen gewählten Organis
mus lokal Rekonfigurationsleistungen realisiert und Produktions- und Verarbeitungsverfahren für biologisches Material verfügbar sind, mit deren Hilfe neues Wissen generiert und dargestellt wer
den kann.
In molekularbiologischen Labors, so eine der Konsequenzen dieser Überlegungen, werden Mäuse nicht nur als ’Modellsysteme für andere Säuger4 analysiert. Als Exemplare einer Kategorie innerhalb der biologischen Systematik erleiden sie im Prozeß ihrer infrastrukturellen ’Disziplinie
rung4 in Laboren einen drastischen Bedeutungs
und körperlichen Identitätswandel. Die Hausmaus wird durch Labormäuse ersetzt, die Produkte und Bestandteile von Laboratopen sind. Als Labor
maus ist ihre natürliche Herkunft gegenüber ihrer Züchtungsbiographie rudimentär. Hergebrachte Klassifikationen werden durch neue, davon radi
kal verschiedene ersetzt. Während die bereits er
wähnte Veränderung der Klassifikationslogik und -praxis im Bereich der bakteriellen Technofakte zunächst als eine Randerscheinung und Nebenfol
ge der Technisierung biologischer Forschung be
handelt werden kann, erscheint die Transforma
tion verschiedener anderer, vor allem höherer Or
ganismen als Modellsysteme die bisherige Klassifi
kationspraxis der Biologie an zentralen Stellen zu untergraben. Aus der einheitlichen A rt der taxono- misch identifizierten Maus, z.B . mus musculus, werden komplexe, autonome Taxonomien gene
riert und mit Technofakten bevölkert. Es handelt sich um reverse Taxonomien, Ordnungen, die das klassische Taxonomieprinzip verkehren: wo tradi
tionell die ’natürliche4 Einheit des Klassifizierten intendiert war, wird Variabilität maximiert. Diese Variationen können nur noch in geschlossenen Sy
stemen von Laboratopen als Bestandteile einer sy