Effect of different mutations in the ATM gene on the cellular response to ionizing radiation
77
0
0
Volltext
(2)
(3)
(4)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(18)
(20)
(21)
(22)
(23)
(24)
(25)
(26)
(27)
(28)
(29)
(30)
(31)
Abbildung
+7
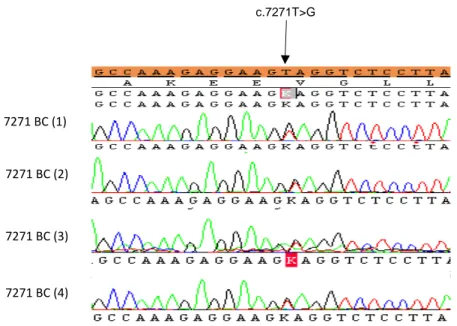
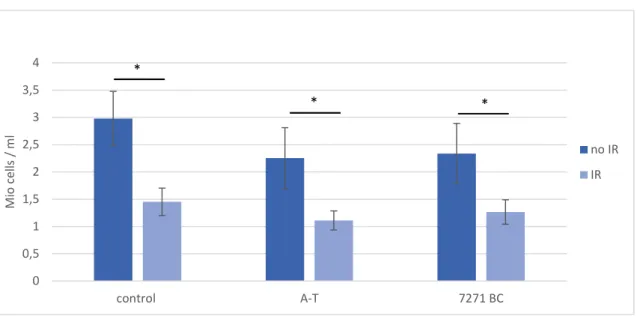
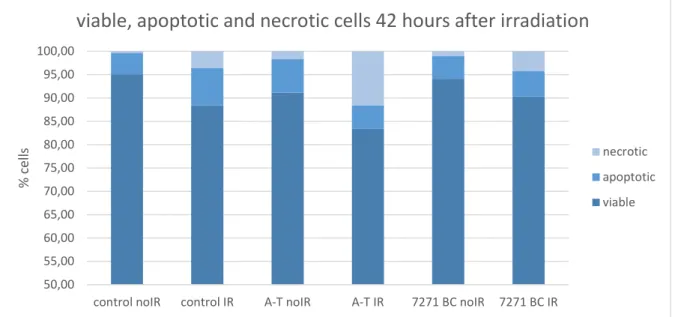
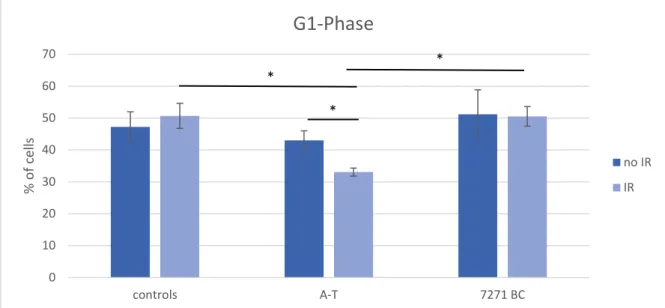


ÄHNLICHE DOKUMENTE
Additionally, within the irradiated samples four different characteristic representatives (without ectoine, 0.2 Gy and 14.07 Gy respectively/ectoine solution, 0.26 Gy and 15.83
Currents, number of primary electrons are measured, the calculation of irradiation doses is based on Hahn et.. All samples are analyzed by
Our real-time PCR experiments confirmed the results of the microarray analysis that OSTL is expressed at lower levels in ALL than in myeloid leukemia, especially CML.. The ALL
To test for a potential role of the demethylase in the DNA damage response, cell cycle analysis experiments and recruitment kinetics of damage response proteins
In my thesis I have proposed and developed a statistical approach that allows study- ing molecular mechanisms of temporal gene expression responses from a time-course
The frequency of detected mutations in the known genes associated with hereditary pancreatic cancer is difficult because patients with known deleterious variants in the
Whereas the Notch2 +/HCS mutation favors pro-osteoclastogenic gene expression, triggering a high bone turnover pathology, the Wnt1 G177C/G177C mutation specifically affects
In the present study I analyzed the signature of miR-16, miR-30b and miR-93 in exosomes derived from plasma of BC patients and compared it with that in DCIS patients and healthy
